

The KRAB zinc finger protein RSL1 regulates sex- and tissue-specific promoter methylation and dynamic hormone-responsive chromatin configuration
- PMID: 22801370
- PMCID: PMC3430191
- DOI: 10.1128/MCB.00615-12
The KRAB zinc finger protein RSL1 regulates sex- and tissue-specific promoter methylation and dynamic hormone-responsive chromatin configuration
Abstract
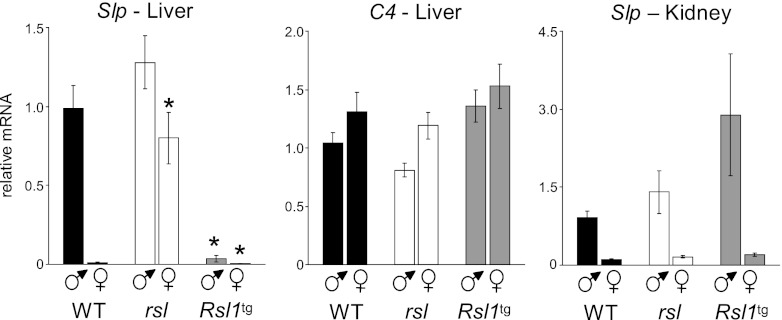
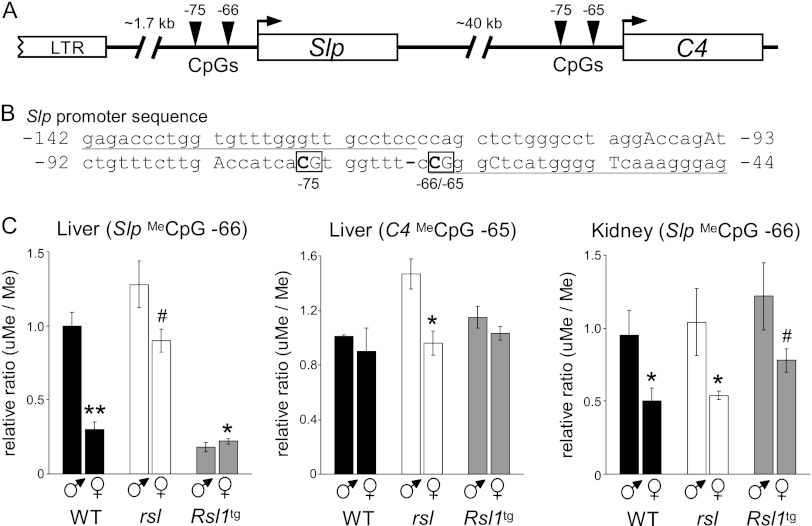
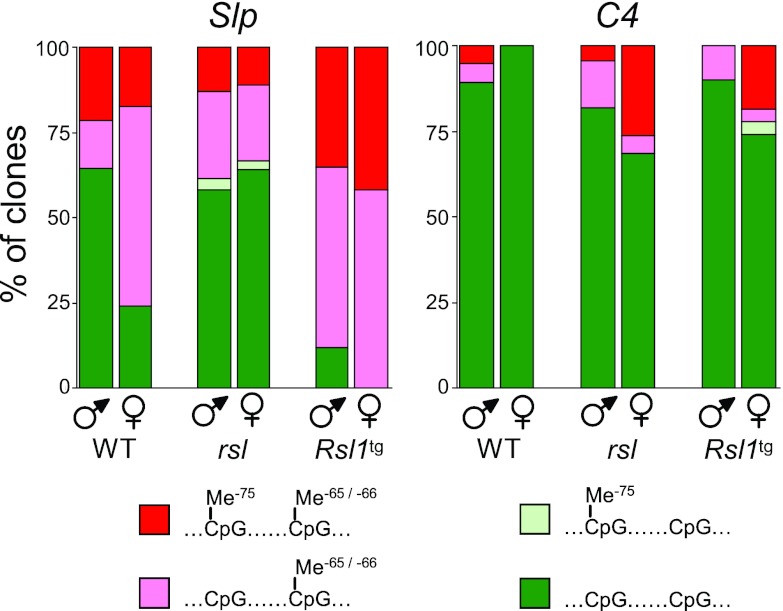
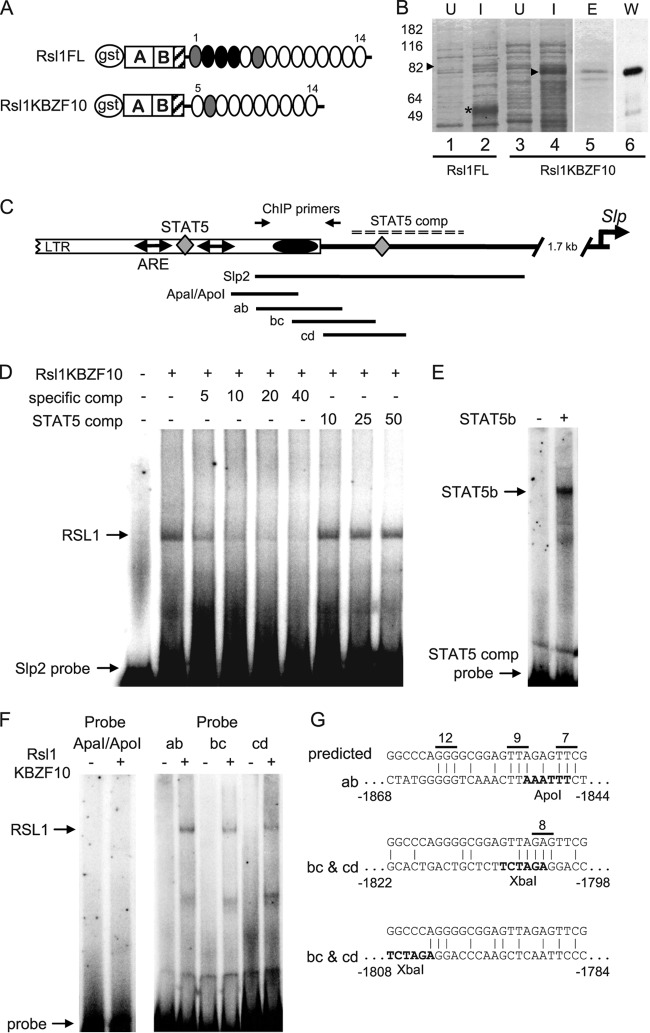
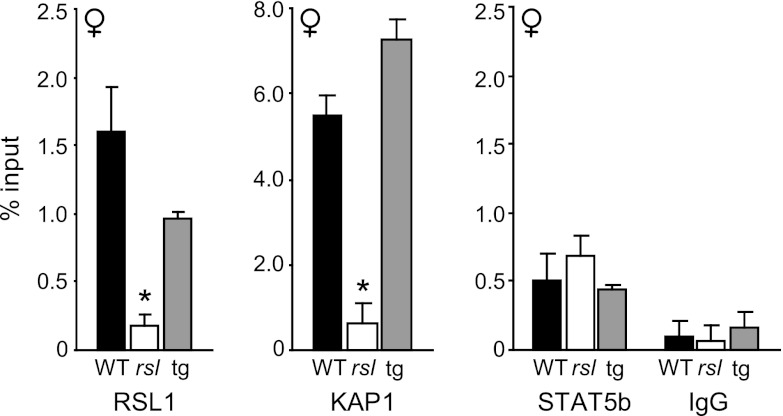
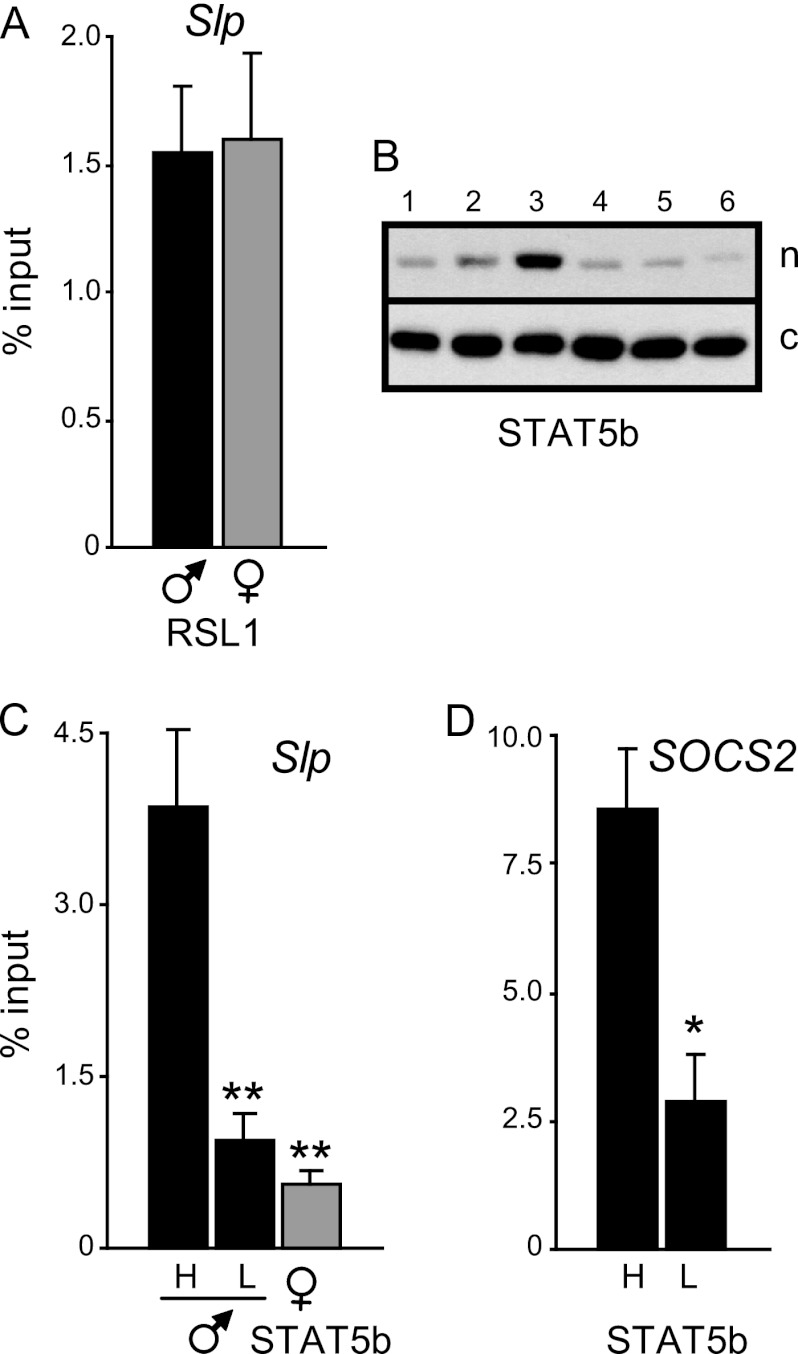
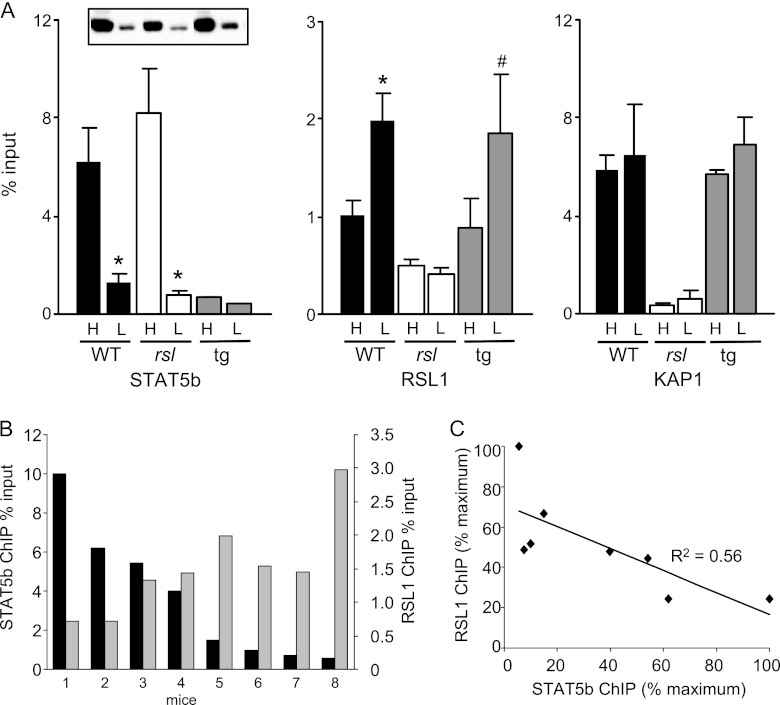
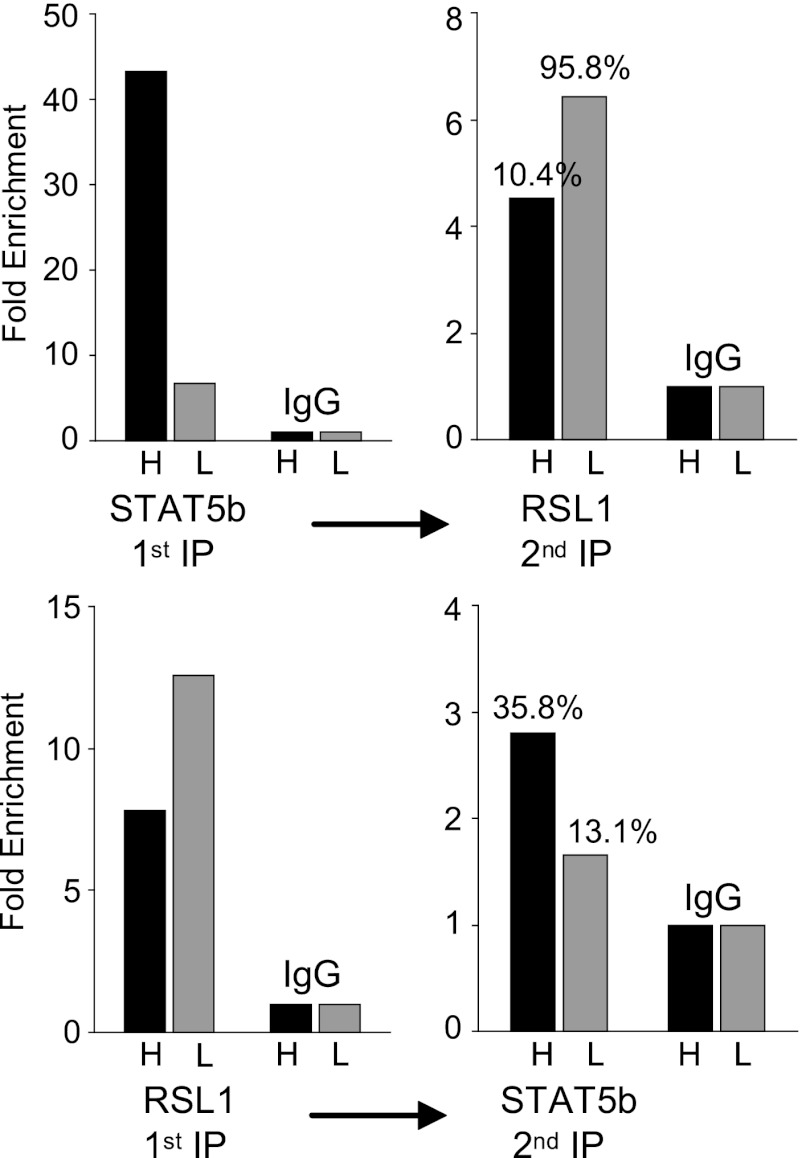
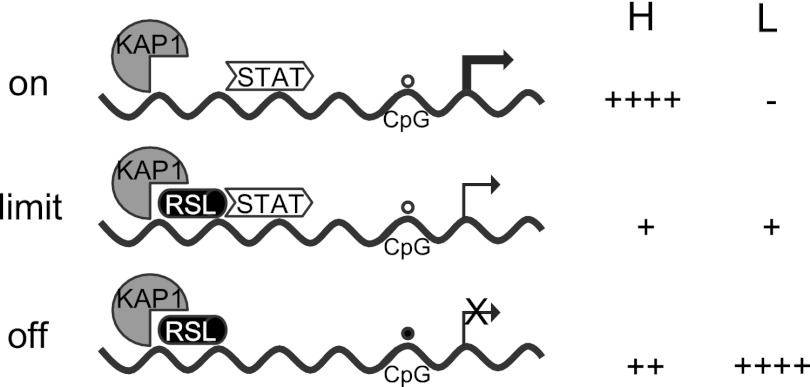
Over 400 Krüppel-associated box zinc finger proteins (KRAB-ZFPs) are encoded in mammalian genomes. While KRAB-ZFPs strongly repress transcription in vitro, little is known about their biological function or gene targets in vivo. Regulator of sex limitation 1 (Rsl1), one of the first KRAB-Zfp genes assigned a physiological role, accentuates sex-biased liver gene expression, most dramatically for mouse sex-limited protein (Slp), which provides an in vivo reporter of KRAB-ZFP function. Slp is induced in males in the liver and kidney by growth hormone (GH) and androgen, respectively. In the liver but not kidney, the Rsl1 genotype correlates with methylation of a CpG dinucleotide in the Slp promoter that is demethylated at puberty. RSL1 binds 2 kb upstream of the Slp promoter, both in vitro and in vivo, within an enhancer containing response elements for STAT5b. Chromatin immunoprecipitation (ChIP) assays demonstrate that RSL1 recruits KAP1/TRIM28, the corepressor for KRAB action in vitro, to this enhancer. Slp induction requires rapid cycling of STAT5b in chromatin. Remarkably, RSL1 simultaneously binds adjacent to STAT5b with a reciprocal binding pattern that limits hormonal response. These experiments demonstrate a surprisingly dynamic interplay between a hormonal activator, STAT5b, and a KRAB-ZFP repressor and provide unique insights into KRAB-ZFP epigenetic mechanisms.
Figures
References
-
- Choi HK, Waxman DJ. 2000. Pulsatility of growth hormone (GH) signalling in liver cells: role of the JAK-STAT5b pathway in GH action. Growth Horm. IGF Res. 10(Suppl B): S1–S8 - PubMed
-
- Chomczynski P, Sacchi N. 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162: 156–159 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous