

Temporal profiling and pulsed SILAC labeling identify novel secreted proteins during ex vivo osteoblast differentiation of human stromal stem cells
- PMID: 22801418
- PMCID: PMC3494153
- DOI: 10.1074/mcp.M111.012138
Temporal profiling and pulsed SILAC labeling identify novel secreted proteins during ex vivo osteoblast differentiation of human stromal stem cells
Abstract
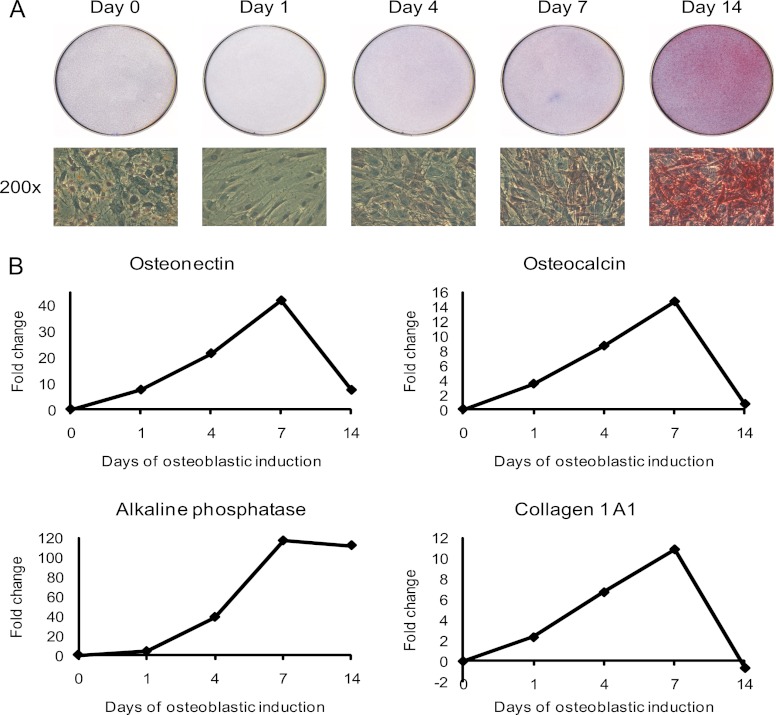
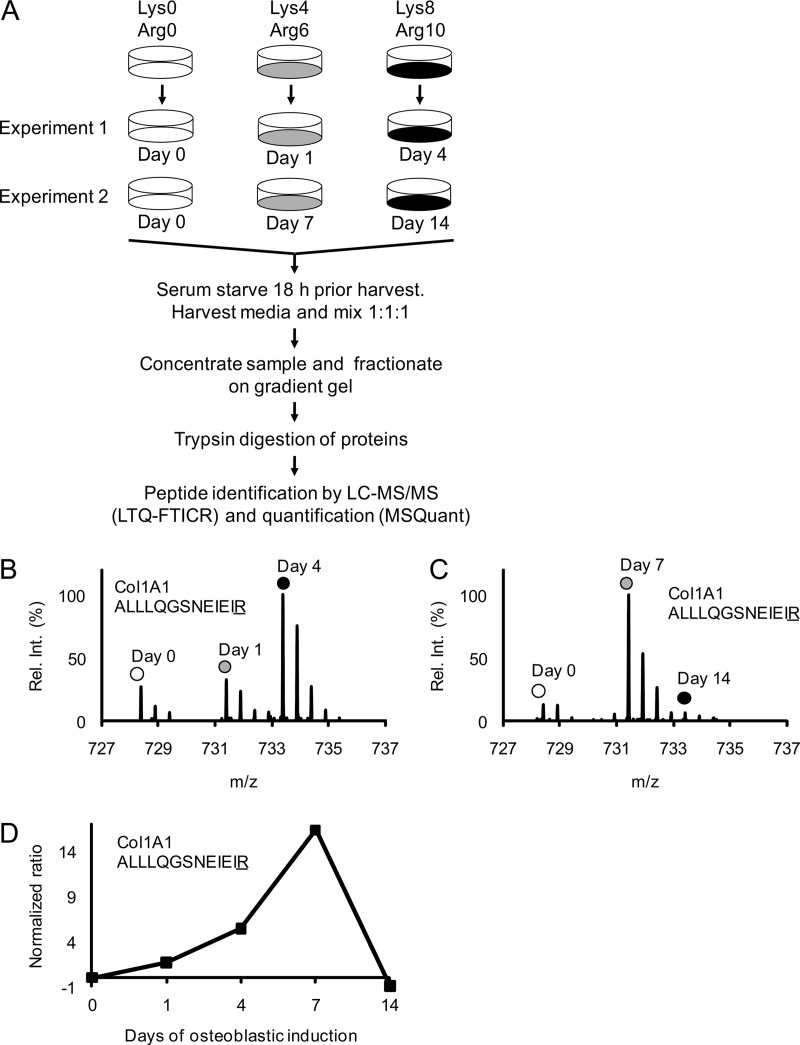
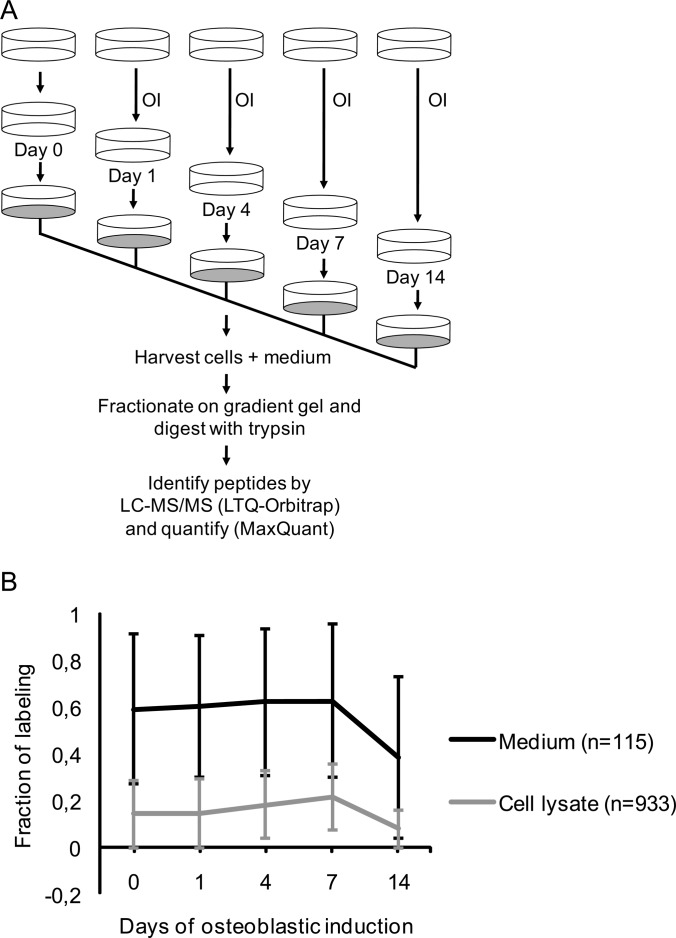
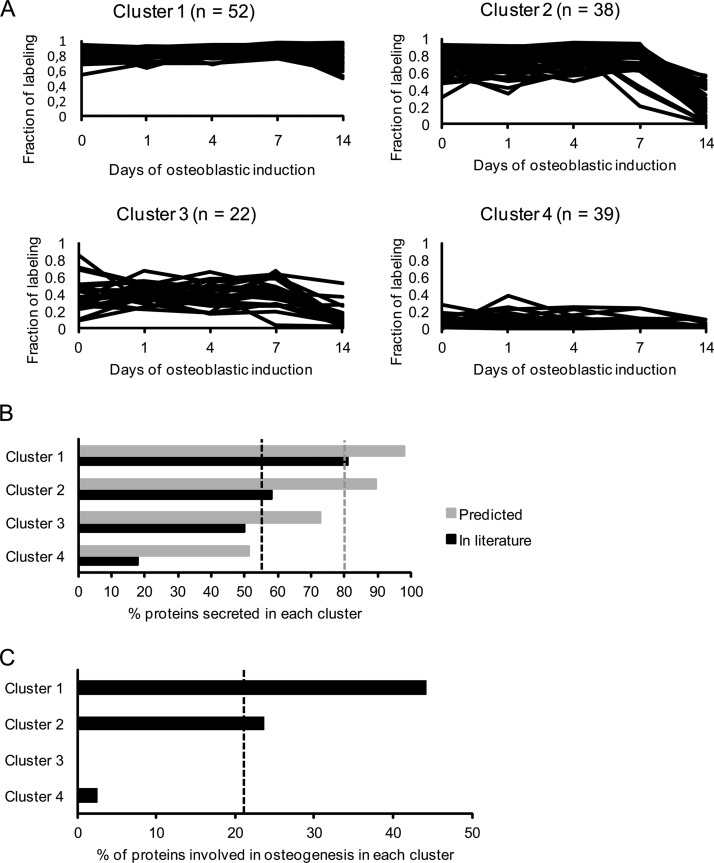
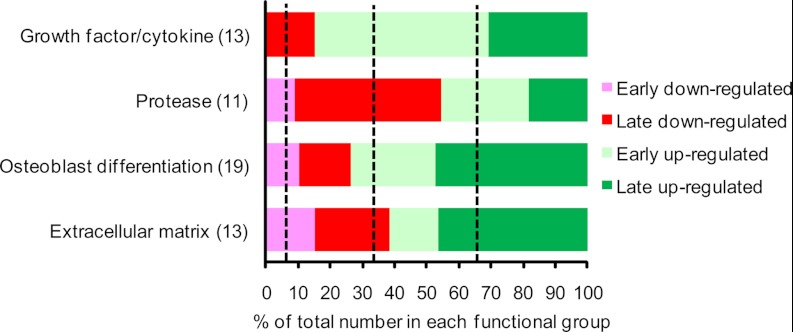
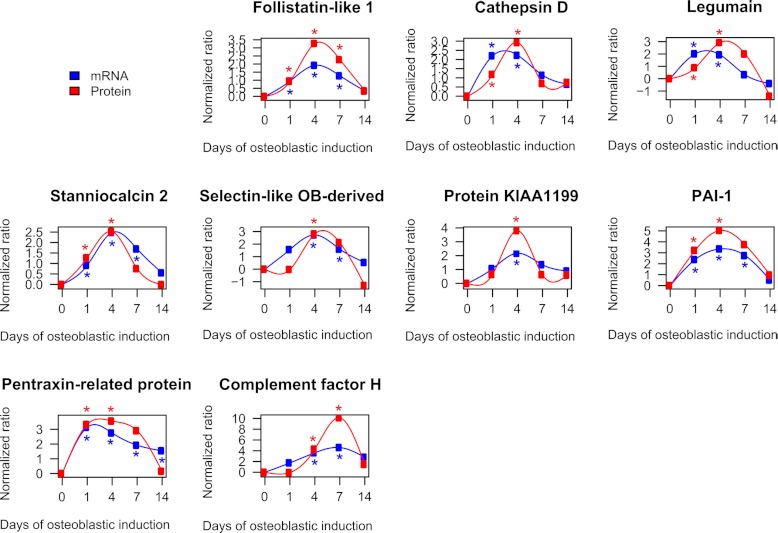
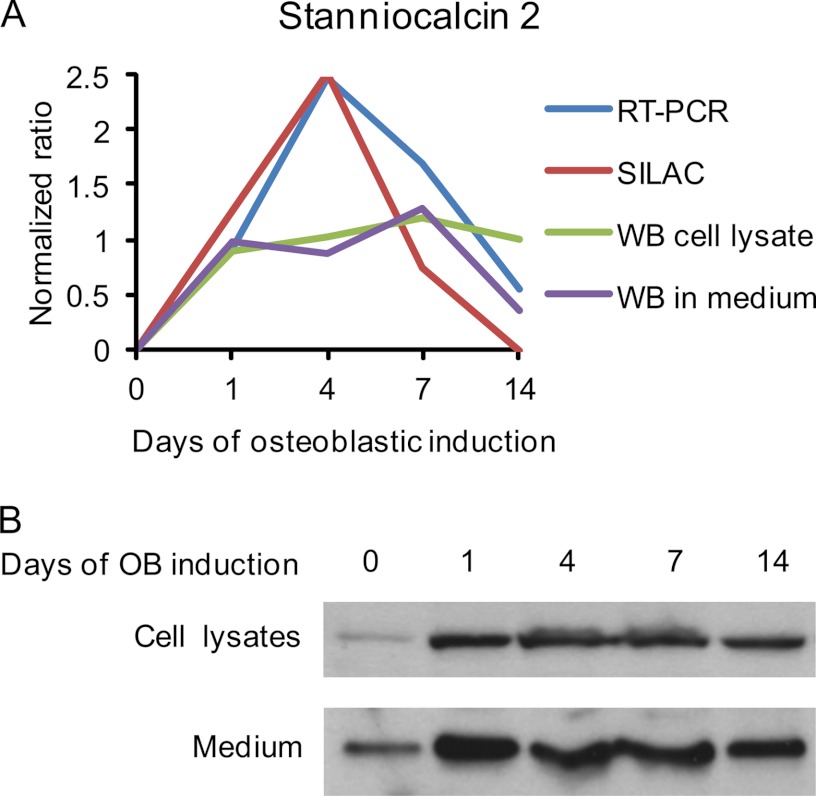
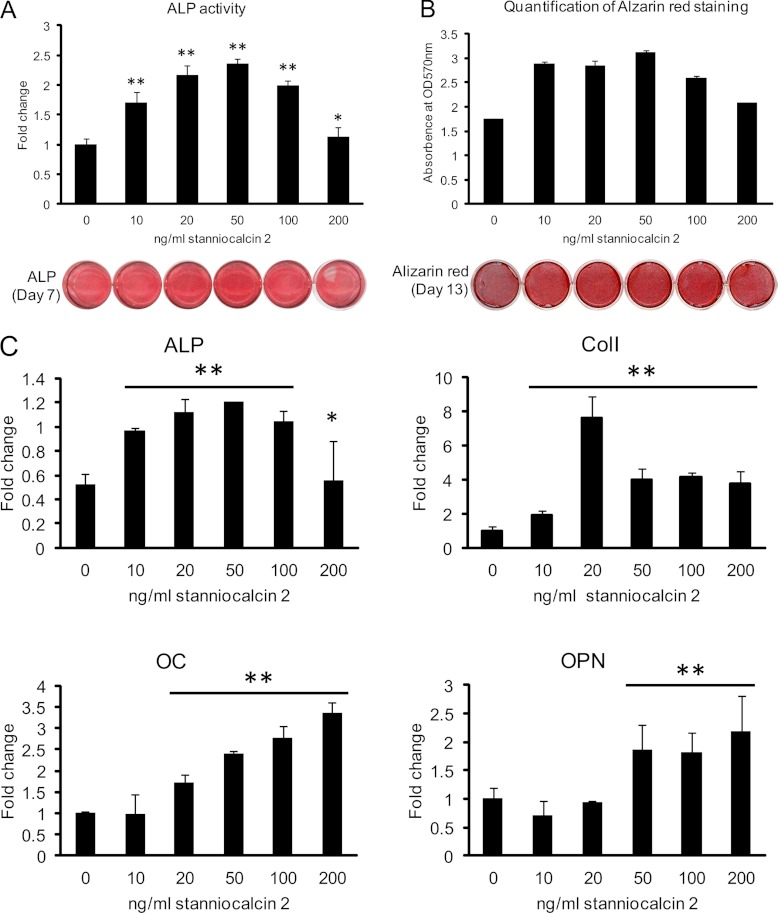
It is well established that bone forming cells (osteoblasts) secrete proteins with autocrine, paracrine, and endocrine function. However, the identity and functional role for the majority of these secreted and differentially expressed proteins during the osteoblast (OB) differentiation process, is not fully established. To address these questions, we quantified the temporal dynamics of the human stromal (mesenchymal, skeletal) stem cell (hMSC) secretome during ex vivo OB differentiation using stable isotope labeling by amino acids in cell culture (SILAC). In addition, we employed pulsed SILAC labeling to distinguish genuine secreted proteins from intracellular contaminants. We identified 466 potentially secreted proteins that were quantified at 5 time-points during 14-days ex vivo OB differentiation including 41 proteins known to be involved in OB functions. Among these, 315 proteins exhibited more than 2-fold up or down-regulation. The pulsed SILAC method revealed a strong correlation between the fraction of isotope labeling and the subset of proteins known to be secreted and involved in OB differentiation. We verified SILAC data using qRT-PCR analysis of 9 identified potential novel regulators of OB differentiation. Furthermore, we studied the biological effects of one of these proteins, the hormone stanniocalcin 2 (STC2) and demonstrated its autocrine effects in enhancing osteoblastic differentiation of hMSC. In conclusion, combining complete and pulsed SILAC labeling facilitated the identification of novel factors produced by hMSC with potential role in OB differentiation. Our study demonstrates that the secretome of osteoblastic cells is more complex than previously reported and supports the emerging evidence that osteoblastic cells secrete proteins with endocrine functions and regulate cellular processes beyond bone formation.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
