

No evidence for inositol 1,4,5-trisphosphate-dependent Ca2+ release in isolated fibers of adult mouse skeletal muscle
- PMID: 22802359
- PMCID: PMC3409103
- DOI: 10.1085/jgp.201110747
No evidence for inositol 1,4,5-trisphosphate-dependent Ca2+ release in isolated fibers of adult mouse skeletal muscle
Abstract
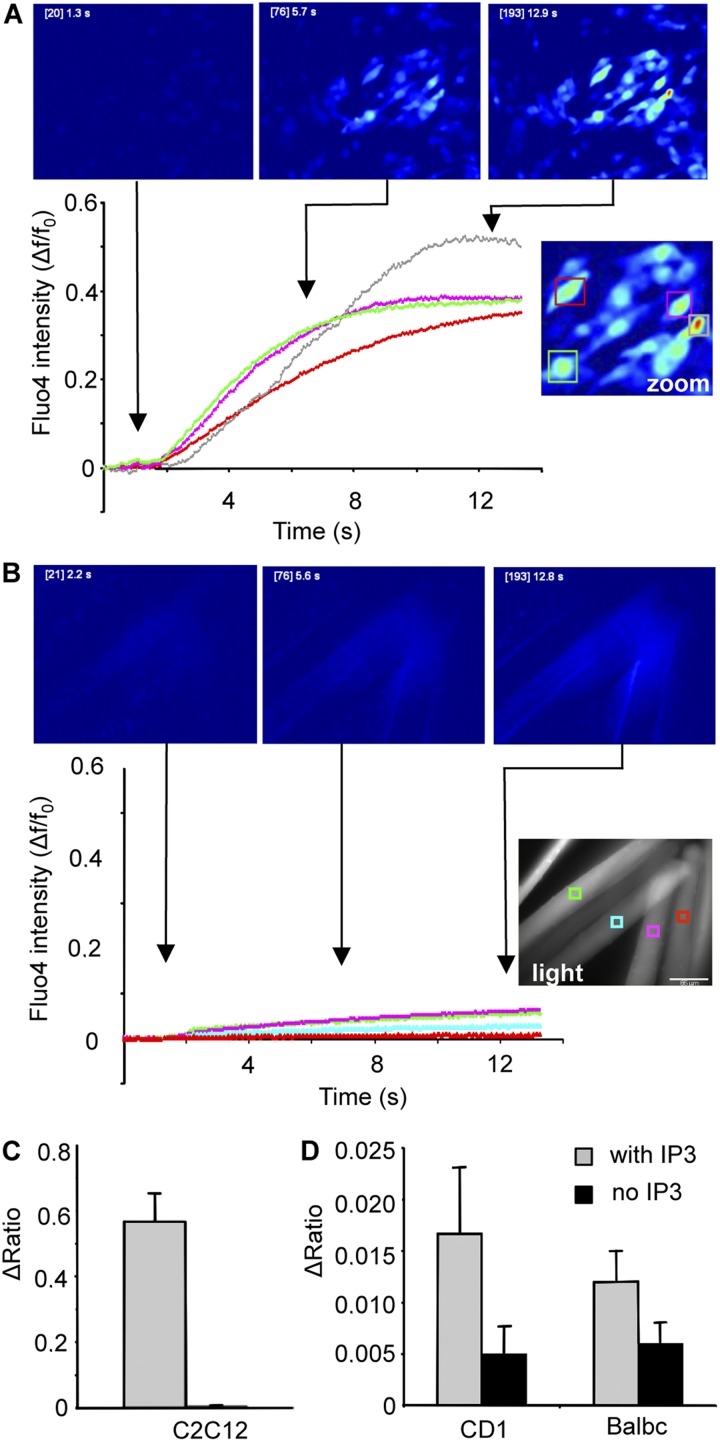
The presence and role of functional inositol 1,4,5-trisphosphate (IP(3)) receptors (IP(3)Rs) in adult skeletal muscle are controversial. The current consensus is that, in adult striated muscle, the relative amount of IP(3)Rs is too low and the kinetics of Ca(2+) release from IP(3)R is too slow compared with ryanodine receptors to contribute to the Ca(2+) transient during excitation-contraction coupling. However, it has been suggested that IP(3)-dependent Ca(2+) release may be involved in signaling cascades leading to regulation of muscle gene expression. We have reinvestigated IP(3)-dependent Ca(2+) release in isolated flexor digitorum brevis (FDB) muscle fibers from adult mice. Although Ca(2+) transients were readily induced in cultured C2C12 muscle cells by (a) UTP stimulation, (b) direct injection of IP(3), or (c) photolysis of membrane-permeant caged IP(3), no statistically significant change in calcium signal was detected in adult FDB fibers. We conclude that the IP(3)-IP(3)R system does not appear to affect global calcium levels in adult mouse skeletal muscle.
Figures


Similar articles
-
Hydroxylated xestospongins block inositol-1,4,5-trisphosphate-induced Ca2+ release and sensitize Ca2+-induced Ca2+ release mediated by ryanodine receptors.Mol Pharmacol. 2006 Feb;69(2):532-8. doi: 10.1124/mol.105.019125. Epub 2005 Oct 25. Mol Pharmacol. 2006. PMID: 16249374
-
IP(3)-dependent, post-tetanic calcium transients induced by electrostimulation of adult skeletal muscle fibers.J Gen Physiol. 2010 Oct;136(4):455-67. doi: 10.1085/jgp.200910397. Epub 2010 Sep 13. J Gen Physiol. 2010. PMID: 20837675 Free PMC article.
-
The contribution of inositol 1,4,5-trisphosphate and ryanodine receptors to agonist-induced Ca(2+) signaling of airway smooth muscle cells.Am J Physiol Lung Cell Mol Physiol. 2009 Aug;297(2):L347-61. doi: 10.1152/ajplung.90559.2008. Epub 2009 May 22. Am J Physiol Lung Cell Mol Physiol. 2009. PMID: 19465516 Free PMC article.
-
Secretory granules in inositol 1,4,5-trisphosphate-dependent Ca2+ signaling in the cytoplasm of neuroendocrine cells.FASEB J. 2010 Mar;24(3):653-64. doi: 10.1096/fj.09-132456. Epub 2009 Oct 16. FASEB J. 2010. PMID: 19837865 Free PMC article. Review.
-
Atrial local Ca2+ signaling and inositol 1,4,5-trisphosphate receptors.Prog Biophys Mol Biol. 2010 Sep;103(1):59-70. doi: 10.1016/j.pbiomolbio.2010.02.002. Epub 2010 Mar 1. Prog Biophys Mol Biol. 2010. PMID: 20193706 Review.
Cited by
-
Type 1 inositol (1,4,5)-trisphosphate receptor activates ryanodine receptor 1 to mediate calcium spark signaling in adult mammalian skeletal muscle.J Biol Chem. 2013 Jan 25;288(4):2103-9. doi: 10.1074/jbc.M112.425975. Epub 2012 Dec 5. J Biol Chem. 2013. PMID: 23223241 Free PMC article.
-
Mitochondrial Calcium Increase Induced by RyR1 and IP3R Channel Activation After Membrane Depolarization Regulates Skeletal Muscle Metabolism.Front Physiol. 2018 Jun 25;9:791. doi: 10.3389/fphys.2018.00791. eCollection 2018. Front Physiol. 2018. PMID: 29988564 Free PMC article.
-
Dynamics of Calcium during In vitro Microspore Embryogenesis and In vivo Microspore Development in Brassica napus and Solanum melongena.Front Plant Sci. 2017 Jul 7;8:1177. doi: 10.3389/fpls.2017.01177. eCollection 2017. Front Plant Sci. 2017. PMID: 28736567 Free PMC article.
-
Angiotensin II modulates mouse skeletal muscle resting conductance to chloride and potassium ions and calcium homeostasis via the AT1 receptor and NADPH oxidase.Am J Physiol Cell Physiol. 2014 Oct 1;307(7):C634-47. doi: 10.1152/ajpcell.00372.2013. Epub 2014 Jul 30. Am J Physiol Cell Physiol. 2014. PMID: 25080489 Free PMC article.
-
Monovalent cationic channel activity in the inner membrane of nuclei from skeletal muscle fibers.Biophys J. 2014 Nov 4;107(9):2027-36. doi: 10.1016/j.bpj.2014.09.030. Biophys J. 2014. PMID: 25418088 Free PMC article.
References
-
- Carrasco M.A., Riveros N., Ríos J., Müller M., Torres F., Pineda J., Lantadilla S., Jaimovich E. 2003. Depolarization-induced slow calcium transients activate early genes in skeletal muscle cells. Am. J. Physiol. Cell Physiol. 284:C1438–C1447 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
