

Serine/threonine acetylation of TGFβ-activated kinase (TAK1) by Yersinia pestis YopJ inhibits innate immune signaling
- PMID: 22802624
- PMCID: PMC3411953
- DOI: 10.1073/pnas.1008203109
Serine/threonine acetylation of TGFβ-activated kinase (TAK1) by Yersinia pestis YopJ inhibits innate immune signaling
Abstract
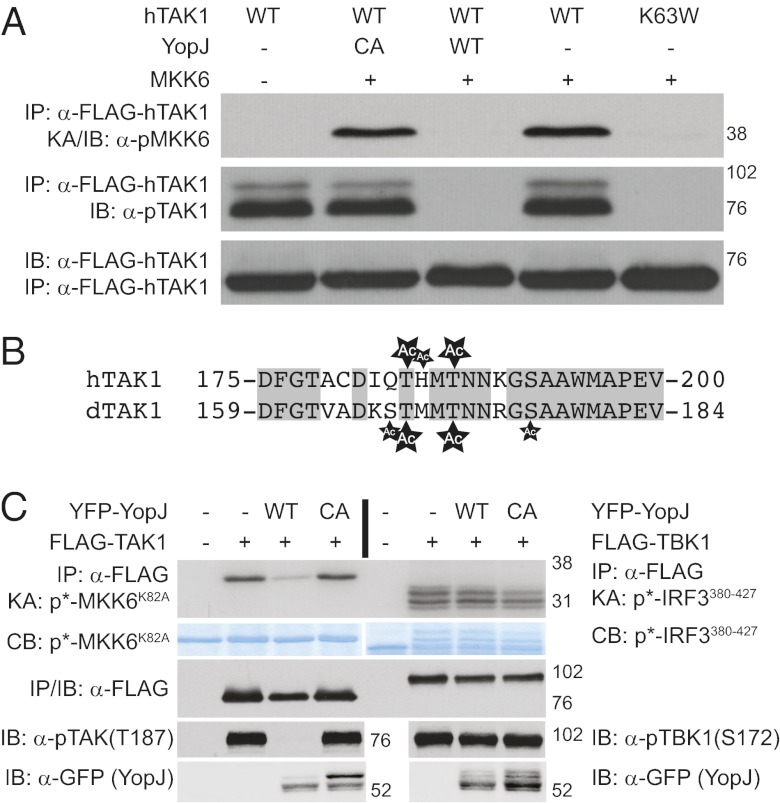
The Gram-negative bacteria Yersinia pestis, causative agent of plague, is extremely virulent. One mechanism contributing to Y. pestis virulence is the presence of a type-three secretion system, which injects effector proteins, Yops, directly into immune cells of the infected host. One of these Yop proteins, YopJ, is proapoptotic and inhibits mammalian NF-κB and MAP-kinase signal transduction pathways. Although the molecular mechanism remained elusive for some time, recent work has shown that YopJ acts as a serine/threonine acetyl-transferase targeting MAP2 kinases. Using Drosophila as a model system, we find that YopJ inhibits one innate immune NF-κB signaling pathway (IMD) but not the other (Toll). In fact, we show YopJ mediated serine/threonine acetylation and inhibition of dTAK1, the critical MAP3 kinase in the IMD pathway. Acetylation of critical serine/threonine residues in the activation loop of Drosophila TAK1 blocks phosphorylation of the protein and subsequent kinase activation. In addition, studies in mammalian cells show similar modification and inhibition of hTAK1. These data present evidence that TAK1 is a target for YopJ-mediated inhibition.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

 marks unmodified full-length IMD,
marks unmodified full-length IMD,  highlights phosphorylated IMD, and
highlights phosphorylated IMD, and  marks the cleaved-IMD products. (C) Phospho-JNK was monitored in whole-cell lysates. Full-length JNK blot serves as a loading control. (D) IKK activity was monitored in S2* cells expressing YopJWT or YopJCA after stimulation with PGN. IKKγ blot serves as an immunoprecipitation control.
marks the cleaved-IMD products. (C) Phospho-JNK was monitored in whole-cell lysates. Full-length JNK blot serves as a loading control. (D) IKK activity was monitored in S2* cells expressing YopJWT or YopJCA after stimulation with PGN. IKKγ blot serves as an immunoprecipitation control.


References
-
- Palmer LE, Hobbie S, Galán JE, Bliska JB. YopJ of Yersinia pseudotuberculosis is required for the inhibition of macrophage TNF-alpha production and downregulation of the MAP kinases p38 and JNK. Mol Microbiol. 1998;27:953–965. - PubMed
-
- Schesser K, et al. The yopJ locus is required for Yersinia-mediated inhibition of NF-kappaB activation and cytokine expression: YopJ contains a eukaryotic SH2-like domain that is essential for its repressive activity. Mol Microbiol. 1998;28:1067–1079. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
