

Social structures depend on innate determinants and chemosensory processing in Drosophila
- PMID: 22802679
- PMCID: PMC3477376
- DOI: 10.1073/pnas.1121252109
Social structures depend on innate determinants and chemosensory processing in Drosophila
Abstract
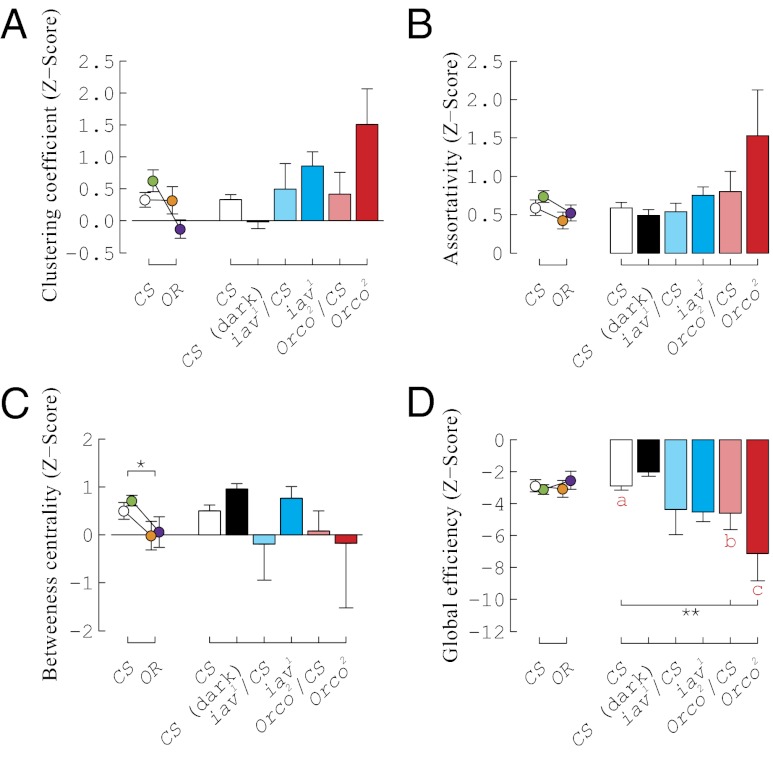
Flies display transient social interactions in groups. However, whether fly-fly interactions are stochastic or structured remains unknown. We hypothesized that groups of flies exhibit patterns of social dynamics that would manifest as nonrandom social interaction networks. To test this, we applied a machine vision system to track the position and orientation of flies in an arena and designed a classifier to detect interactions between pairs of flies. We show that the vinegar fly, Drosophila melanogaster, forms nonrandom social interaction networks, distinct from virtual network controls (constructed from the intersections of individual locomotor trajectories). In addition, the formation of interaction networks depends on chemosensory cues. Gustatory mutants form networks that cannot be distinguished from their virtual network controls. Olfactory mutants form networks that are greatly disrupted compared with control flies. Different wild-type strains form social interaction networks with quantitatively different properties, suggesting that the genes that influence this network phenotype vary across and within wild-type populations. We have established a paradigm for studying social behaviors at a group level in Drosophila and expect that a genetic dissection of this phenomenon will identify conserved molecular mechanisms of social organization in other species.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Shorrocks B. Drosophila. London: Ginn; 1972.
-
- Sokolowski MB. Social interactions in “simple” model systems. Neuron. 2010;65:780–794. - PubMed
-
- Wertheim B, van Baalen EJ, Dicke M, Vet LE. Pheromone-mediated aggregation in nonsocial arthropods: An evolutionary ecological perspective. Annu Rev Entomol. 2005;50:321–346. - PubMed
-
- Mery F, et al. Public versus personal information for mate copying in an invertebrate. Curr Biol. 2009;19:730–734. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
