

Indoleamine 2,3-dioxygenase (IDO) induced by Leishmania infection of human dendritic cells
- PMID: 22803643
- PMCID: PMC3572781
- DOI: 10.1111/j.1365-3024.2012.01380.x
Indoleamine 2,3-dioxygenase (IDO) induced by Leishmania infection of human dendritic cells
Abstract
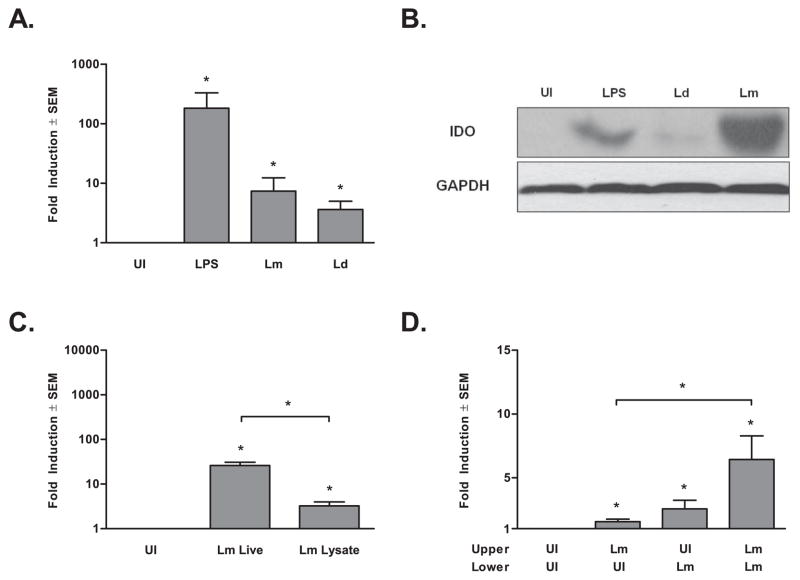
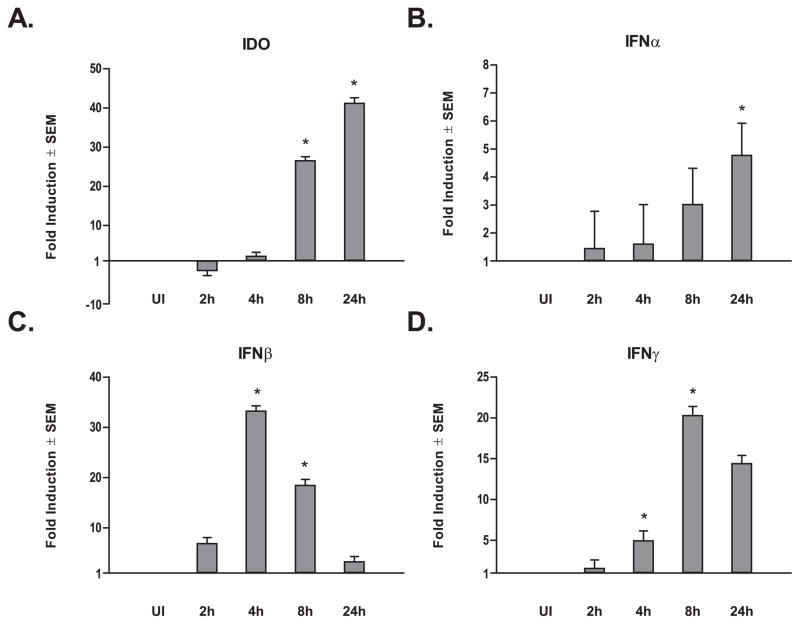
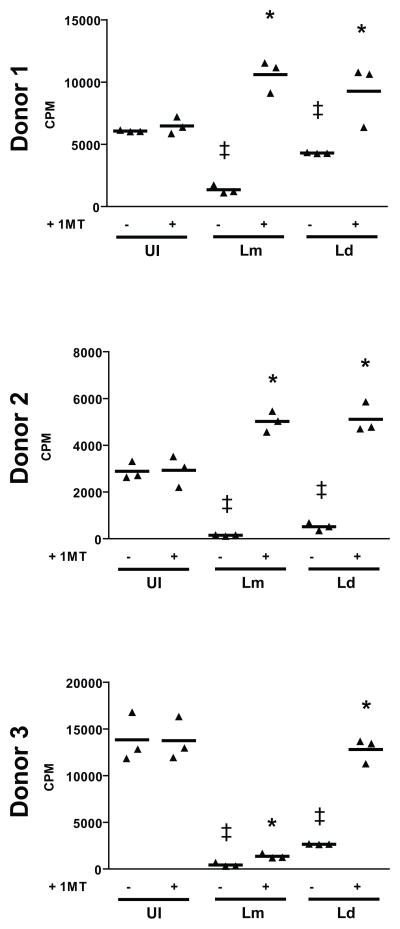
Dendritic cells (DC) play a pivotal role in regulating immunity, establishing immunologically privileged tissue microenvironments and maintaining homoeostasis. It is becoming increasingly clear that one key mechanism that mediates many DC functions is production of the immunomodulatory enzyme indoleamine 2,3-dioxygenase (IDO). For pathogens that cause chronic infection, exploitation of host DCs is a solution to establish and persist within a host. Leishmania parasites cause a range of clinical manifestations, all involving chronic infection, and are proficient at avoiding immune responses. We demonstrate here that infection of human myeloid-derived DC with L. major and L. donovani induces IDO expression using a mechanism that involves autocrine or paracrine stimulation with a DC-secreted factor. Leishmania-induced IDO suppresses allogeneic and tetanus toxoid-specific lymphocyte proliferation, an inhibition that is reversed with the IDO inhibitor, 1-methyl tryptophan (1-MT). Furthermore, IDO expression by human DC does not require live Leishmania infection, as parasite lysates also up-regulate IDO mRNA production. Our data suggest that one mechanism Leishmania parasites utilize to circumvent immune clearance may be to promote the induction of IDO among host DC within the infection microenvironment.
© 2012 Blackwell Publishing Ltd.
Figures

Similar articles
-
Leishmania major attenuates host immunity by stimulating local indoleamine 2,3-dioxygenase expression.J Infect Dis. 2011 Mar 1;203(5):715-25. doi: 10.1093/infdis/jiq095. Epub 2011 Jan 31. J Infect Dis. 2011. PMID: 21282196 Free PMC article.
-
Indoleamine 2,3-dioxygenase production by human dendritic cells results in the inhibition of T cell proliferation.J Immunol. 2000 Apr 1;164(7):3596-9. doi: 10.4049/jimmunol.164.7.3596. J Immunol. 2000. PMID: 10725715
-
Lysophosphatidylcholine exacerbates Leishmania major-dendritic cell infection through interleukin-10 and a burst in arginase1 and indoleamine 2,3-dioxygenase activities.Int Immunopharmacol. 2015 Mar;25(1):1-9. doi: 10.1016/j.intimp.2015.01.006. Epub 2015 Jan 16. Int Immunopharmacol. 2015. PMID: 25601495
-
IDO-expressing regulatory dendritic cells in cancer and chronic infection.J Mol Med (Berl). 2008 Feb;86(2):145-60. doi: 10.1007/s00109-007-0262-6. Epub 2007 Sep 18. J Mol Med (Berl). 2008. PMID: 17876564 Review.
-
The modulation of enzyme indoleamine 2,3-dioxygenase from dendritic cells for the treatment of type 1 diabetes mellitus.Drug Des Devel Ther. 2017 Jul 24;11:2171-2178. doi: 10.2147/DDDT.S135367. eCollection 2017. Drug Des Devel Ther. 2017. PMID: 28769554 Free PMC article. Review.
Cited by
-
Splenic Transcriptional Responses in Severe Visceral Leishmaniasis: Impaired Leukocyte Chemotaxis and Cell Cycle Arrest.Front Immunol. 2021 Nov 5;12:716314. doi: 10.3389/fimmu.2021.716314. eCollection 2021. Front Immunol. 2021. PMID: 34804009 Free PMC article.
-
Drug analog inhibition of indoleamine 2,3-dioxygenase (IDO) activity modifies pattern recognition receptor expression and proinflammatory cytokine responses early during influenza virus infection.J Leukoc Biol. 2014 Sep;96(3):447-52. doi: 10.1189/jlb.3AB0114-046RR. Epub 2014 May 5. J Leukoc Biol. 2014. PMID: 24799604 Free PMC article.
-
Human dendritic cells exhibit a pronounced type I IFN signature following Leishmania major infection that is required for IL-12 induction.J Immunol. 2014 Jun 15;192(12):5863-72. doi: 10.4049/jimmunol.1203230. Epub 2014 May 7. J Immunol. 2014. PMID: 24808365 Free PMC article.
-
Early reduction in PD-L1 expression predicts faster treatment response in human cutaneous leishmaniasis.J Clin Invest. 2021 Nov 15;131(22):e142765. doi: 10.1172/JCI142765. J Clin Invest. 2021. PMID: 34609968 Free PMC article.
-
Transcriptional profiling of the spleen in progressive visceral leishmaniasis reveals mixed expression of type 1 and type 2 cytokine-responsive genes.BMC Immunol. 2014 Nov 26;15:38. doi: 10.1186/s12865-014-0038-z. BMC Immunol. 2014. PMID: 25424735 Free PMC article.
References
-
- Maizels R. Regulation of the immune system in metazoan parasite infections. Novartis Found Symp. 2007;281:192–204. discussion -9. - PubMed
-
- Sacks D, Sher A. Evasion of innate immunity by parasitic protozoa. Nat Immunol. 2002 Nov;3(11):1041–7. - PubMed
-
- McDowell MA, Sacks DL. Inhibition of host cell signal transduction by Leishmania: observations relevant to the selective impairment of IL-12 responses. Curr Opin Microbiol. 1999;2(4):438–43. - PubMed
-
- Peters N, Sacks D. Immune privilege in sites of chronic infection: Leishmania and regulatory T cells. Immunol Rev. 2006 Oct;213:159–79. - PubMed
-
- Choi CM, Lerner EA. Leishmaniasis as an emerging infection. J Investig Dermatol Symp Proc. 2001 Dec;6(3):175–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
