

New insights into the troubles of aneuploidy
- PMID: 22804579
- PMCID: PMC3919630
- DOI: 10.1146/annurev-cellbio-101011-155807
New insights into the troubles of aneuploidy
Abstract
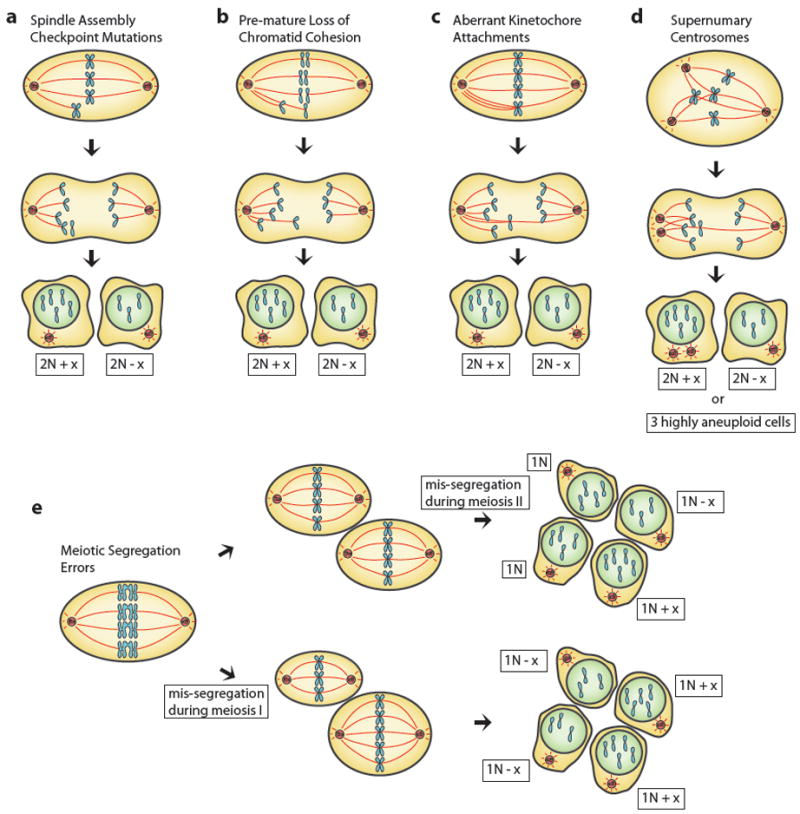
Deviation from a balanced genome by either gain or loss of entire chromosomes is generally tolerated poorly in all eukaryotic systems studied to date. Errors in mitotic or meiotic cell division lead to aneuploidy, which places a burden of additional or insufficient gene products from the missegregated chromosomes on the daughter cells. The burden of aneuploidy often manifests itself as impaired fitness of individual cells and whole organisms, in which abnormal development is also characteristic. However, most human cancers, noted for their rapid growth, also display various levels of aneuploidy. Here we discuss the detrimental, potentially beneficial, and sometimes puzzling effects of aneuploidy on cellular and organismal fitness and tissue function as well as its role in diseases such as cancer and neurodegeneration.
Figures





References
-
- Aguilar MJ, Kamoshita S, Landing BH, Boder E, Sedgwick RP. Pathological observations in ataxia-telangiectasia. A report of five cases. J Neuropathol Exp Neurol. 1968;27:659–76. - PubMed
-
- Albertson DG, Collins C, McCormick F, Gray JW. Chromosome aberrations in solid tumors. Nat Genet. 2003;34:369–76. - PubMed
-
- Baker DJ, Jeganathan KB, Cameron JD, Thompson M, Juneja S, et al. BubR1 insifficiency causes early onset of aging-associated phenotypes and infertility in mice. Nat Genet. 2004;36:744–9. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
