

Erythrocytic pyruvate kinase mutations causing hemolytic anemia, osteosclerosis, and secondary hemochromatosis in dogs
- PMID: 22805166
- PMCID: PMC3650904
- DOI: 10.1111/j.1939-1676.2012.00958.x
Erythrocytic pyruvate kinase mutations causing hemolytic anemia, osteosclerosis, and secondary hemochromatosis in dogs
Abstract
Background: Erythrocytic pyruvate kinase (PK) deficiency, first documented in Basenjis, is the most common inherited erythroenzymopathy in dogs.
Objectives: To report 3 new breed-specific PK-LR gene mutations and a retrospective survey of PK mutations in as mall and selected group of Beagles and West Highland White Terriers (WHWT).
Animals: Labrador Retrievers (2 siblings, 5 unrelated), Pugs (2 siblings, 1 unrelated), Beagles (39 anemic, 29 other),WHWTs (22 anemic, 226 nonanemic), Cairn Terrier (n = 1).
Methods: Exons of the PK-LR gene were sequenced from genomic DNA of young dogs (<2 years) with persistent highly regenerative hemolytic anemia.
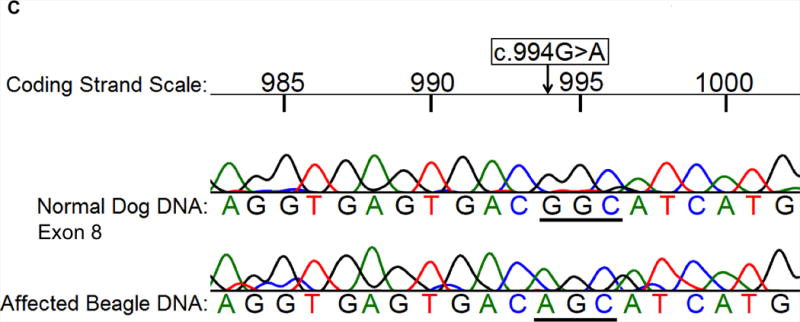
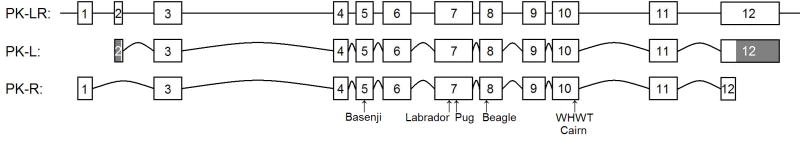
Results: A nonsense mutation (c.799C>T) resulting in a premature stop codon was identified in anemic Labrador Retriever siblings that had osteosclerosis, high serum ferritin concentrations, and severe hepatic secondary hemochromatosis. Anemic Pug and Beagle revealed 2 different missense mutations (c.848T>C, c.994G>A, respectively) resulting in intolerable amino acid changes to protein structure and enzyme function. Breed-specific mutation tests were developed. Among the biased group of 248 WHWTs, 9% and 35% were homozygous (affected) and heterozygous, respectively, for the previously described mutation (mutant allele frequency 0.26). A PK-deficient Cairn Terrier had the same insertion mutation as the affected WHWTs. Of the selected group of 68 Beagles, 35% were PK-deficient and 3% were carriers (0.37).
Conclusions and clinical importance: Erythrocytic PK deficiency is caused by different mutations in different dog breeds and causes chronic severe hemolytic anemia, hemosiderosis, and secondary hemochromatosis because of chronic hemolysis and, an as yet unexplained osteosclerosis. The newly developed breed-specific mutation assays simplify the diagnosis of PK deficiency.
Figures





Similar articles
-
Determination of erythrocyte pyruvate kinase deficiency in Basenjis with chronic hemolytic anemia.J Am Vet Med Assoc. 1991 May 15;198(10):1755-61. J Am Vet Med Assoc. 1991. PMID: 2071475
-
Identification of a 6 base pair insertion in West Highland White Terriers with erythrocyte pyruvate kinase deficiency.Am J Vet Res. 1999 Sep;60(9):1169-72. Am J Vet Res. 1999. PMID: 10490091
-
Genetic test for pyruvate kinase deficiency of Basenjis.J Am Vet Med Assoc. 1995 Oct 1;207(7):918-21. J Am Vet Med Assoc. 1995. PMID: 7559024
-
Molecular basis of erythroenzymopathies associated with hereditary hemolytic anemia: tabulation of mutant enzymes.Am J Hematol. 1996 Feb;51(2):122-32. doi: 10.1002/(SICI)1096-8652(199602)51:2<122::AID-AJH5>3.0.CO;2-#. Am J Hematol. 1996. PMID: 8579052 Review.
-
Red cell enzymopathies as a model of inborn errors of metabolism.Southeast Asian J Trop Med Public Health. 1995;26 Suppl 1:112-9. Southeast Asian J Trop Med Public Health. 1995. PMID: 8629088 Review.
Cited by
-
Genetic Panel Screening of Nearly 100 Mutations Reveals New Insights into the Breed Distribution of Risk Variants for Canine Hereditary Disorders.PLoS One. 2016 Aug 15;11(8):e0161005. doi: 10.1371/journal.pone.0161005. eCollection 2016. PLoS One. 2016. PMID: 27525650 Free PMC article.
-
Web resource on available DNA variant tests for hereditary diseases and genetic predispositions in dogs and cats: An Update.Hum Genet. 2021 Nov;140(11):1505-1515. doi: 10.1007/s00439-021-02256-5. Epub 2021 Feb 6. Hum Genet. 2021. PMID: 33547946
-
Normal glucose metabolism in carnivores overlaps with diabetes pathology in non-carnivores.Front Endocrinol (Lausanne). 2013 Dec 3;4:188. doi: 10.3389/fendo.2013.00188. Front Endocrinol (Lausanne). 2013. PMID: 24348462 Free PMC article. Review.
-
Erythrocyte pyruvate kinase deficiency mutation identified in multiple breeds of domestic cats.BMC Vet Res. 2012 Oct 30;8:207. doi: 10.1186/1746-6148-8-207. BMC Vet Res. 2012. PMID: 23110753 Free PMC article.
-
Erythrocyte pyruvate kinase deficiency in three West Highland white terriers in Ireland and the UK.Ir Vet J. 2013 Jul 10;66(1):12. doi: 10.1186/2046-0481-66-12. Ir Vet J. 2013. PMID: 23842571 Free PMC article.
References
-
- Harvey JW. Erythrocyte biochemistry. In: Weiss DJ, Wardrop KJ, editors. Schalm’s Veterinary Hematology. 6. Ames, IA, USA: Wiley-Blackwell; 2010. pp. 131–135.
-
- Giger U. Hereditary erythrocyte enzyme abnormalities. In: Weiss DJ, Wardrop KJ, editors. Schalm’s Veterinary Hematology. 6. Ames, IA, USA: Wiley-Blackwell; 2010. pp. 179–186.
-
- Giger U, Noble NA. Determination of eythrocyte pyruvate kinase deficiency in Basenjis with chronic hemolytic anemias. J Am Vet Med A. 1991;198:1755–1761. - PubMed
-
- Whitney KM, Lothrop CD. Genetic test for pyruvate kinase deficiency of Basenjis. J Am Vet Med A. 1995;207:918– 921. - PubMed
-
- Whitney KM, Goodman SA, Bailey EM, Lothrop CD., Jr The molecular basis of canine pyruvate kinase deficiency. Exp Hematol. 1994;22:866–874. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
