

GSH or palmitate preserves mitochondrial energetic/redox balance, preventing mechanical dysfunction in metabolically challenged myocytes/hearts from type 2 diabetic mice
- PMID: 22807033
- PMCID: PMC3501888
- DOI: 10.2337/db12-0072
GSH or palmitate preserves mitochondrial energetic/redox balance, preventing mechanical dysfunction in metabolically challenged myocytes/hearts from type 2 diabetic mice
Abstract
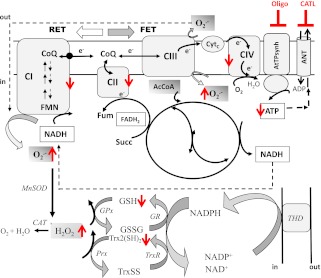
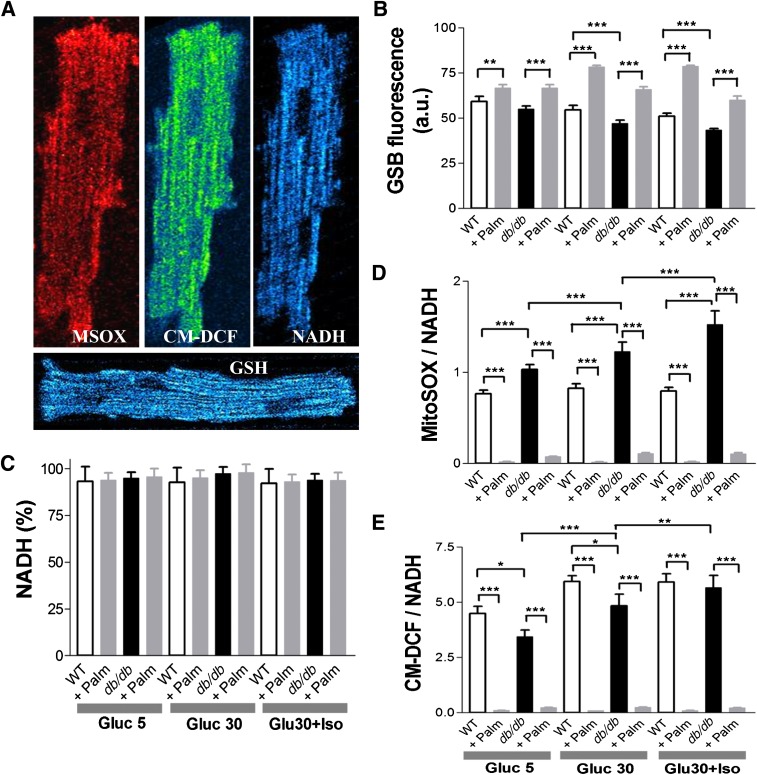
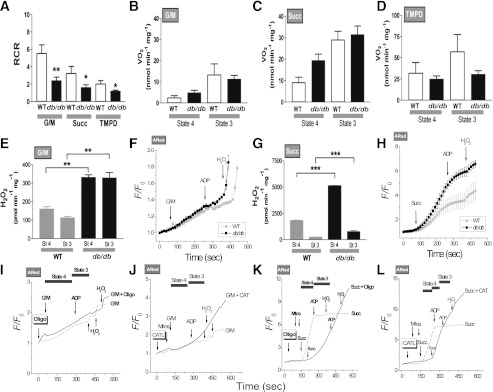
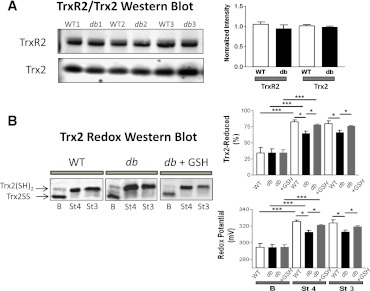
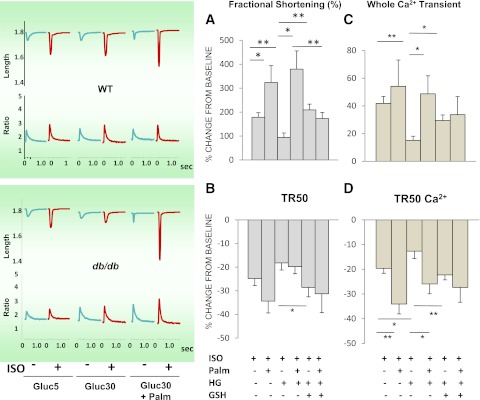
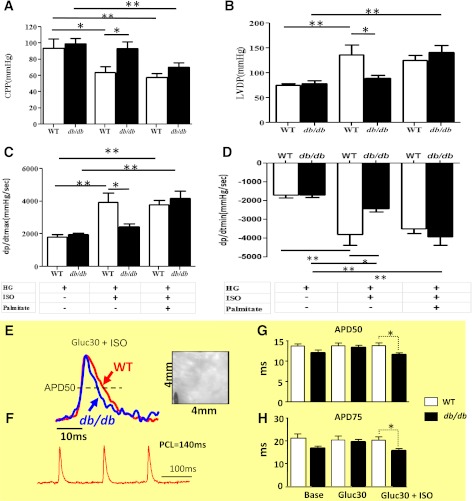
In type 2 diabetes, hyperglycemia and increased sympathetic drive may alter mitochondria energetic/redox properties, decreasing the organelle's functionality. These perturbations may prompt or sustain basal low-cardiac performance and limited exercise capacity. Yet the precise steps involved in this mitochondrial failure remain elusive. Here, we have identified dysfunctional mitochondrial respiration with substrates of complex I, II, and IV and lowered thioredoxin-2/glutathione (GSH) pools as the main processes accounting for impaired state 4→3 energetic transition shown by mitochondria from hearts of type 2 diabetic db/db mice upon challenge with high glucose (HG) and the β-agonist isoproterenol (ISO). By mimicking clinically relevant conditions in type 2 diabetic patients, this regimen triggers a major overflow of reactive oxygen species (ROS) from mitochondria that directly perturbs cardiac electro-contraction coupling, ultimately leading to heart dysfunction. Exogenous GSH or, even more so, the fatty acid palmitate rescues basal and β-stimulated function in db/db myocyte/heart preparations exposed to HG/ISO. This occurs because both interventions provide the reducing equivalents necessary to counter mitochondrial ROS outburst and energetic failure. Thus, in the presence of poor glycemic control, the diabetic patient's inability to cope with increased cardiac work demand largely stems from mitochondrial redox/energetic disarrangements that mutually influence each other, leading to myocyte or whole-heart mechanical dysfunction.
Figures
References
-
- Bell DSH. Heart failure: the frequent, forgotten, and often fatal complication of diabetes. Diabetes Care 2003;26:2433–2441 - PubMed
-
- Laakso M. Hyperglycemia and cardiovascular disease in type 2 diabetes. Diabetes 1999;48:937–942 - PubMed
-
- Cohen-Solal A, Beauvais F, Logeart D. Heart failure and diabetes mellitus: epidemiology and management of an alarming association. J Card Fail 2008;14:615–625 - PubMed
-
- Bruce DG, Chisholm DJ, Storlien LH, Kraegen EW, Smythe GA. The effects of sympathetic nervous system activation and psychological stress on glucose metabolism and blood pressure in subjects with type 2 (non-insulin-dependent) diabetes mellitus. Diabetologia 1992;35:835–843 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
