

Characterization of a proteasome and TAP-independent presentation of intracellular epitopes by HLA-B27 molecules
- PMID: 22807446
- PMCID: PMC3436287
- DOI: 10.1074/jbc.M112.384339
Characterization of a proteasome and TAP-independent presentation of intracellular epitopes by HLA-B27 molecules
Abstract
Nascent HLA-class I molecules are stabilized by proteasome-derived peptides in the ER and the new complexes proceed to the cell surface through the post-ER vesicles. It has been shown, however, that less stable complexes can exchange peptides in the Trans Golgi Network (TGN). HLA-B27 are the most studied HLA-class I molecules due to their association with Ankylosing Spondylitis (AS). Chimeric proteins driven by TAT of HIV have been exploited by us to deliver viral epitopes, whose cross-presentation by the HLA-B27 molecules was proteasome and TAP-independent and not restricted to Antigen-Presenting Cells (APC). Here, using these chimeric proteins as epitope suppliers, we compared with each other and with the HLA-A2 molecules, the two HLA-B*2705 and B*2709 alleles differing at residue 116 (D116H) and differentially associated with AS. We found that the antigen presentation by the two HLA-B27 molecules was proteasome-, TAP-, and APC-independent whereas the presentation by the HLA-A2 molecules required proteasome, TAP and professional APC. Assuming that such difference could be due to the unpaired, highly reactive Cys-67 distinguishing the HLA-B27 molecules, C67S mutants in HLA-B*2705 and B*2709 and V67C mutant in HLA-A*0201 were also analyzed. The results showed that this mutation did not influence the HLA-A2-restricted antigen presentation while it drastically affected the HLA-B27-restricted presentation with, however, remarkable differences between B*2705 and B*2709. The data, together with the occurrence on the cell surface of unfolded molecules in the case of C67S-B*2705 mutant but not in that of C67S-B*2709 mutant, indicates that Cys-67 has a more critical role in stabilizing the B*2705 rather than the B*2709 complexes.
Figures



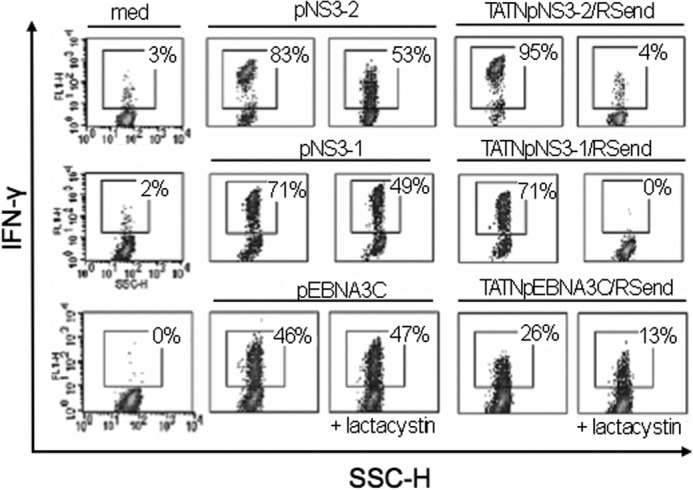
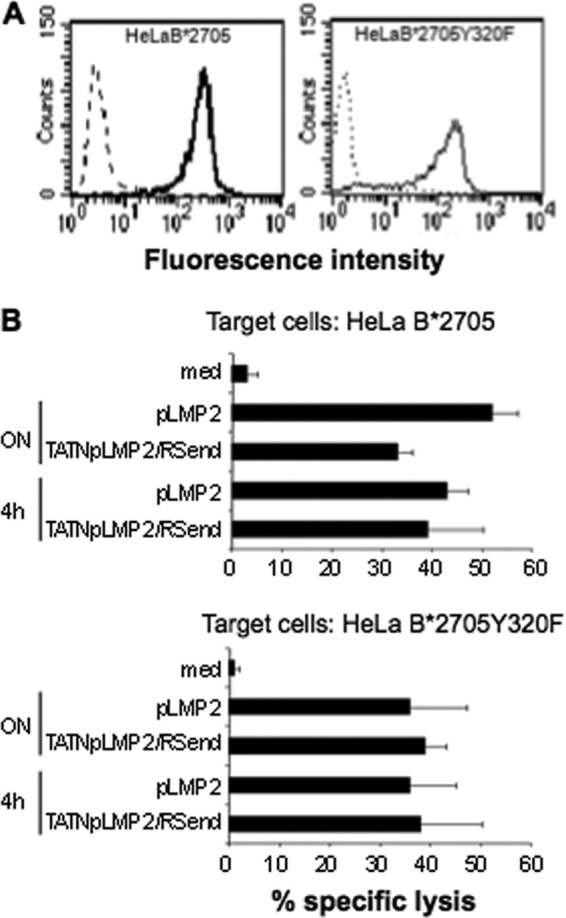




References
-
- Nurzia E., Panimolle F., Cauli A., Mathieu A., Magnacca A., Paladini F., Sorrentino R., Fiorillo M. T. (2010) CD8+ T-cell mediated self-reactivity in HLA-B27 context as a consequence of dual peptide conformation. Clin. Immunol. 135, 476–482 - PubMed
-
- Mathieu A., Paladini F., Vacca A., Cauli A., Fiorillo M. T., Sorrentino R. (2009) The interplay between the geographic distribution of HLA-B27 alleles and their role in infectious and autoimmune diseases: a unifying hypothesis. Autoimmun. Rev. 8, 420–425 - PubMed
-
- Blanco-Gelaz M. A., López-Vázquez A., García-Fernández S., Martínez-Borra J., González S., López-Larrea C. (2001) Genetic variability, molecular evolution, and geographic diversity of HLA-B27. Hum. Immunol. 62, 1042–1050 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
