

Regulation of DNA replication within the immunoglobulin heavy-chain locus during B cell commitment
- PMID: 22807655
- PMCID: PMC3393677
- DOI: 10.1371/journal.pbio.1001360
Regulation of DNA replication within the immunoglobulin heavy-chain locus during B cell commitment
Abstract
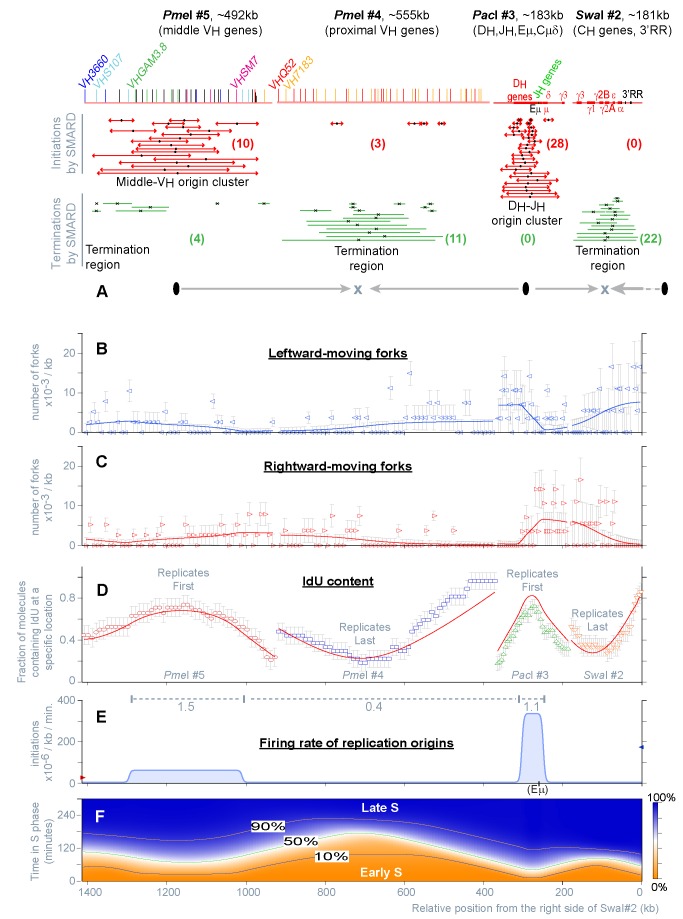
The temporal order of replication of mammalian chromosomes appears to be linked to their functional organization, but the process that establishes and modifies this order during cell differentiation remains largely unknown. Here, we studied how the replication of the Igh locus initiates, progresses, and terminates in bone marrow pro-B cells undergoing B cell commitment. We show that many aspects of DNA replication can be quantitatively explained by a mechanism involving the stochastic firing of origins (across the S phase and the Igh locus) and extensive variations in their firing rate (along the locus). The firing rate of origins shows a high degree of coordination across Igh domains that span tens to hundreds of kilobases, a phenomenon not observed in simple eukaryotes. Differences in domain sizes and firing rates determine the temporal order of replication. During B cell commitment, the expression of the B-cell-specific factor Pax5 sharply alters the temporal order of replication by modifying the rate of origin firing within various Igh domains (particularly those containing Pax5 binding sites). We propose that, within the Igh C(H)-3'RR domain, Pax5 is responsible for both establishing and maintaining high rates of origin firing, mostly by controlling events downstream of the assembly of pre-replication complexes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Norio P, Kosiyatrakul S, Yang Q, Guan Z, Brown N. M, et al. Progressive activation of DNA replication initiation in large domains of the immunoglobulin heavy chain locus during B cell development. Mol Cell. 2005;20:575–587. - PubMed
-
- Kitsberg D, Selig S, Keshet I, Cedar H. Replication structure of the human beta-globin gene domain. Nature. 1993;366:588–590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
