

A trans-species missense SNP in Amhr2 is associated with sex determination in the tiger pufferfish, Takifugu rubripes (fugu)
- PMID: 22807687
- PMCID: PMC3395601
- DOI: 10.1371/journal.pgen.1002798
A trans-species missense SNP in Amhr2 is associated with sex determination in the tiger pufferfish, Takifugu rubripes (fugu)
Abstract
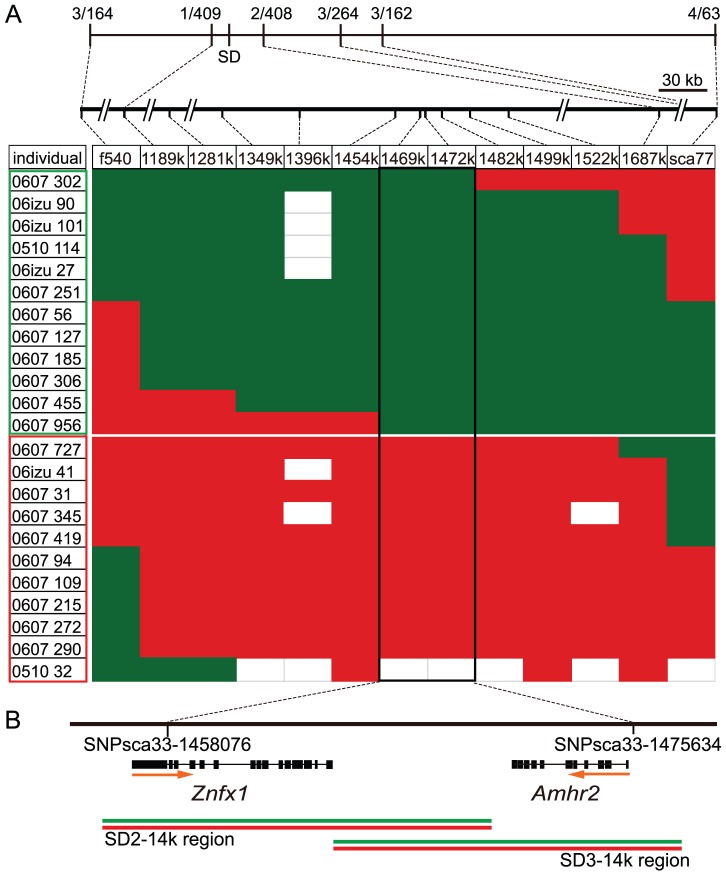
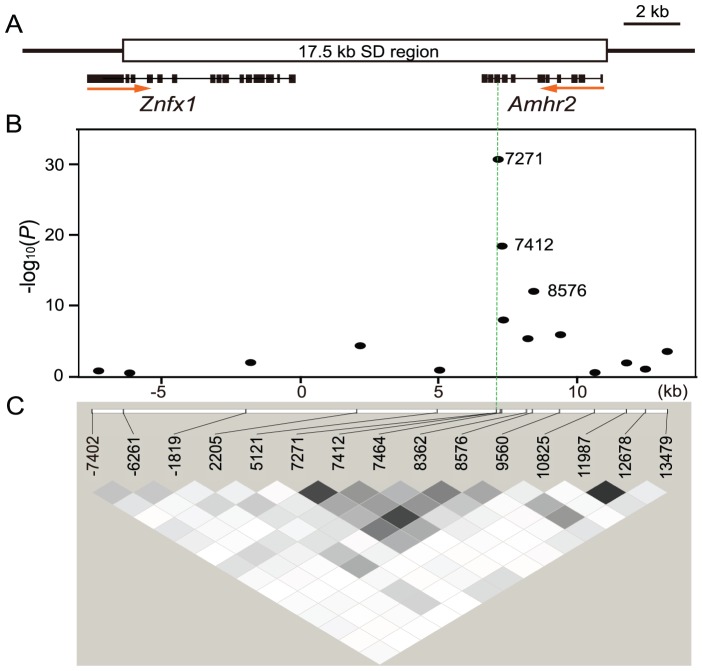
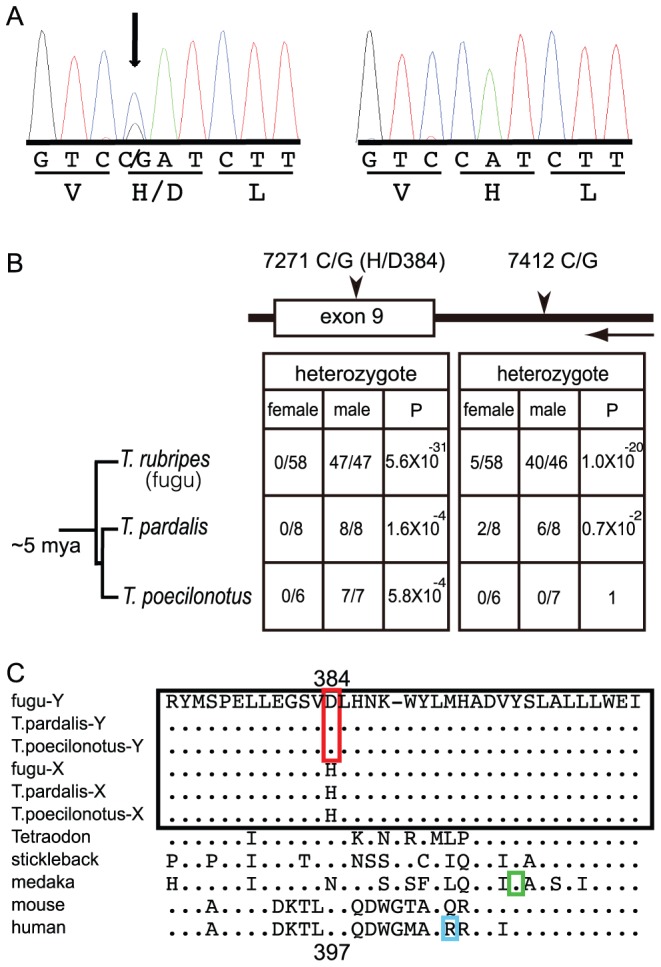
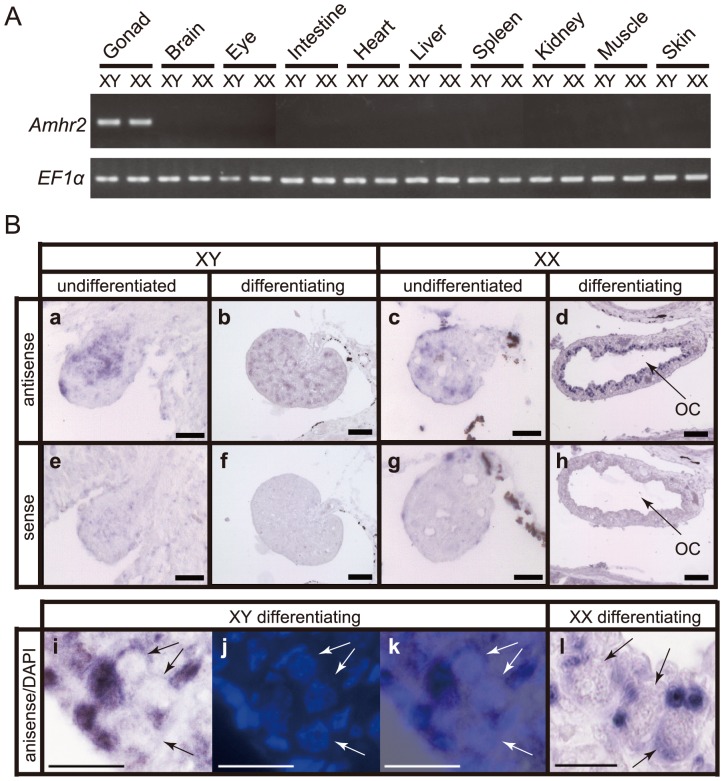
Heterogametic sex chromosomes have evolved independently in various lineages of vertebrates. Such sex chromosome pairs often contain nonrecombining regions, with one of the chromosomes harboring a master sex-determining (SD) gene. It is hypothesized that these sex chromosomes evolved from a pair of autosomes that diverged after acquiring the SD gene. By linkage and association mapping of the SD locus in fugu (Takifugu rubripes), we show that a SNP (C/G) in the anti-Müllerian hormone receptor type II (Amhr2) gene is the only polymorphism associated with phenotypic sex. This SNP changes an amino acid (His/Asp384) in the kinase domain. While females are homozygous (His/His384), males are heterozygous. Sex in fugu is most likely determined by a combination of the two alleles of Amhr2. Consistent with this model, the medaka hotei mutant carrying a substitution in the kinase domain of Amhr2 causes a female phenotype. The association of the Amhr2 SNP with phenotypic sex is conserved in two other species of Takifugu but not in Tetraodon. The fugu SD locus shows no sign of recombination suppression between X and Y chromosomes. Thus, fugu sex chromosomes represent an unusual example of proto-sex chromosomes. Such undifferentiated X-Y chromosomes may be more common in vertebrates than previously thought.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
A sex-linked SNP mutation in amhr2 is responsible for male differentiation in obscure puffer (Takifugu obscurus).Mol Biol Rep. 2021 Aug;48(8):6035-6046. doi: 10.1007/s11033-021-06606-4. Epub 2021 Aug 3. Mol Biol Rep. 2021. PMID: 34341900
-
Identification of the sex-determining locus in grass puffer (Takifugu niphobles) provides evidence for sex-chromosome turnover in a subset of Takifugu species.PLoS One. 2018 Jan 2;13(1):e0190635. doi: 10.1371/journal.pone.0190635. eCollection 2018. PLoS One. 2018. PMID: 29293639 Free PMC article.
-
The sex-determining locus in the tiger pufferfish, Takifugu rubripes.Genetics. 2007 Apr;175(4):2039-42. doi: 10.1534/genetics.106.069278. Epub 2007 Feb 7. Genetics. 2007. PMID: 17287528 Free PMC article.
-
Novel sex-determining genes in fish and sex chromosome evolution.Dev Dyn. 2013 Apr;242(4):339-53. doi: 10.1002/dvdy.23927. Epub 2013 Mar 1. Dev Dyn. 2013. PMID: 23335327 Review.
-
Unusual genomic structure: melanocortin receptors in Fugu.Ann N Y Acad Sci. 2005 Apr;1040:460-3. doi: 10.1196/annals.1327.090. Ann N Y Acad Sci. 2005. PMID: 15891089 Review.
Cited by
-
Preferential Mapping of Sex-Biased Differentially-Expressed Genes of Larvae to the Sex-Determining Region of Flathead Grey Mullet (Mugil cephalus).Front Genet. 2020 Aug 21;11:839. doi: 10.3389/fgene.2020.00839. eCollection 2020. Front Genet. 2020. PMID: 32973865 Free PMC article.
-
Oryzias curvinotus in Sanya Does Not Contain the Male Sex-Determining Gene dmy.Animals (Basel). 2021 May 6;11(5):1327. doi: 10.3390/ani11051327. Animals (Basel). 2021. PMID: 34066583 Free PMC article.
-
Genome assemblies for Chromidotilapia guntheri (Teleostei: Cichlidae) identify a novel candidate gene for vertebrate sex determination, RIN3.Front Genet. 2024 Aug 16;15:1447628. doi: 10.3389/fgene.2024.1447628. eCollection 2024. Front Genet. 2024. PMID: 39221227 Free PMC article.
-
Roles of anti-Müllerian hormone and its duplicates in sex determination and germ cell proliferation of Nile tilapia.Genetics. 2022 Mar 3;220(3):iyab237. doi: 10.1093/genetics/iyab237. Genetics. 2022. PMID: 35100374 Free PMC article.
-
The unusual rainbow trout sex determination gene hijacked the canonical vertebrate gonadal differentiation pathway.Proc Natl Acad Sci U S A. 2018 Dec 11;115(50):12781-12786. doi: 10.1073/pnas.1803826115. Epub 2018 Nov 21. Proc Natl Acad Sci U S A. 2018. PMID: 30463951 Free PMC article.
References
-
- Ohno S. Sex chromosomes and sex-linked genes. Berlin, New York etc.: Springer-Verlag; 1967. 192
-
- Bergero R, Charlesworth D. The evolution of restricted recombination in sex chromosomes. Trends Ecol Evol. 2009;24:94–102. - PubMed
-
- Sinclair AH, Berta P, Palmer MS, Hawkins JR, Griffiths BL, et al. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature. 1990;346:240–244. - PubMed
-
- Smith CA, Roeszler KN, Ohnesorg T, Cummins DM, Farlie PG, et al. The avian Z-linked gene DMRT1 is required for male sex determination in the chicken. Nature. 2009;461:267–271. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
