

Developmental gene expression profiling along the tonotopic axis of the mouse cochlea
- PMID: 22808246
- PMCID: PMC3395647
- DOI: 10.1371/journal.pone.0040735
Developmental gene expression profiling along the tonotopic axis of the mouse cochlea
Abstract
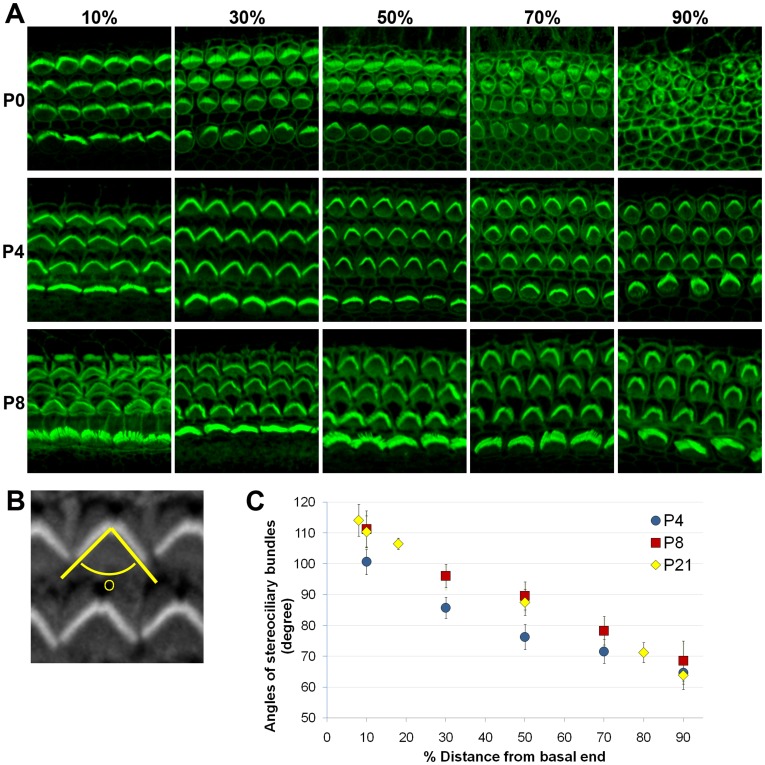
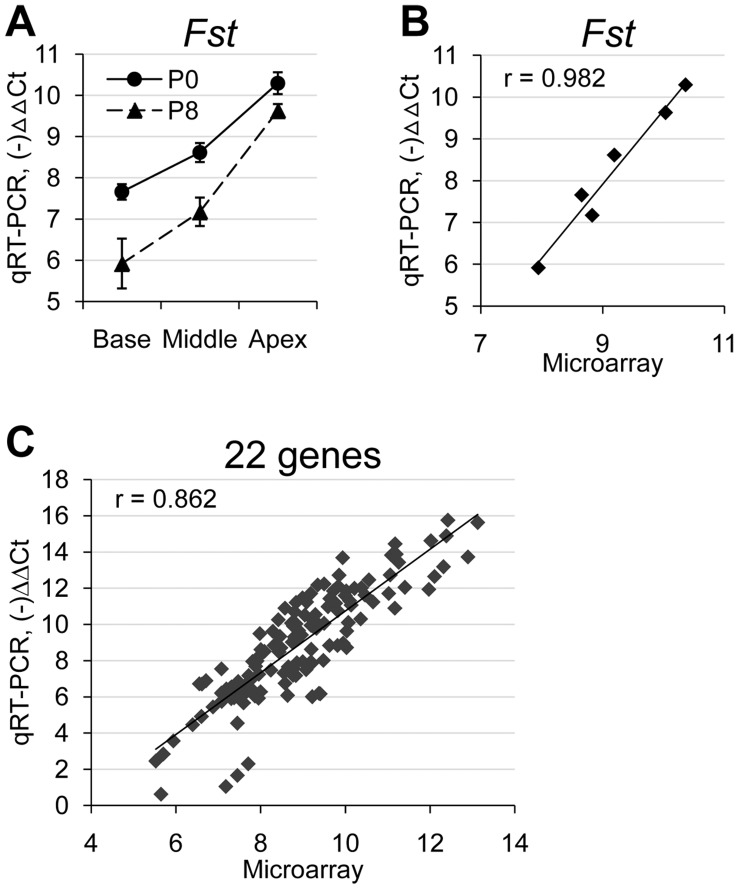
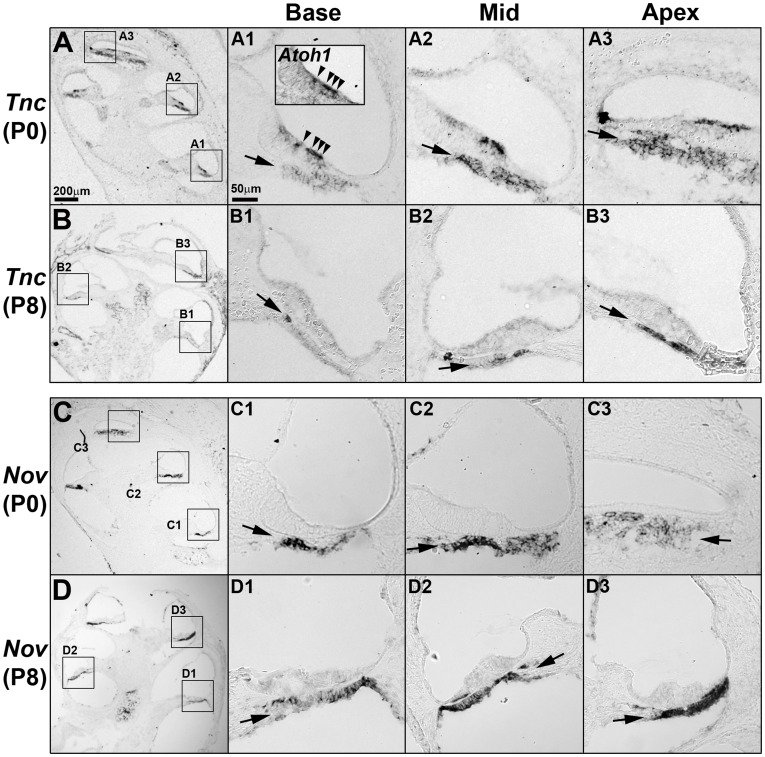
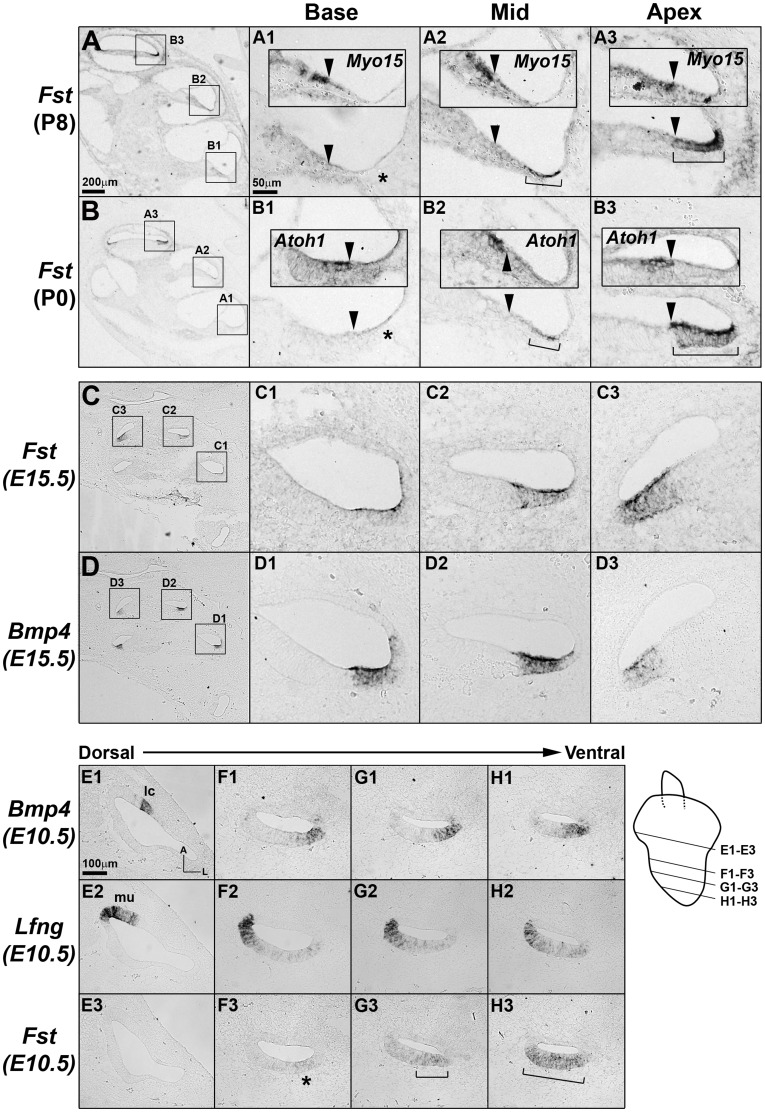
The mammalian cochlear duct is tonotopically organized such that the basal cochlea is tuned to high frequency sounds and the apical cochlea to low frequency sounds. In an effort to understand how this tonotopic organization is established, we searched for genes that are differentially expressed along the tonotopic axis during neonatal development. Cochlear tissues dissected from P0 and P8 mice were divided into three equal pieces, representing the base, middle and apex, and gene expression profiles were determined using the microarray technique. The gene expression profiles were grouped according to changes in expression levels along the tonotopic axis as well as changes during neonatal development. The classified groups were further analyzed by functional annotation clustering analysis to determine whether genes associated with specific biological function or processes are particularly enriched in each group. These analyses identified several candidate genes that may be involved in cochlear development and acquisition of tonotopy. We examined the expression domains for a few candidate genes in the developing mouse cochlea. Tnc (tenacin C) and Nov (nephroblastoma overexpressed gene) are expressed in the basilar membrane, with increased expression toward the apex, which may contribute to graded changes in the structure of the basilar membrane along the tonotopic axis. In addition, Fst (Follistatin), an antagonist of TGF-β/BMP signaling, is expressed in the lesser epithelial ridge and at gradually higher levels towards the apex. The graded expression pattern of Fst is established at the time of cochlear specification and maintained throughout embryonic and postnatal development, suggesting its possible role in the organization of tonotopy. Our data will provide a good resource for investigating the developmental mechanisms of the mammalian cochlea including the acquisition of tonotopy.
Conflict of interest statement
Figures
References
-
- Davis RL. Gradients of neurotrophins, ion channels, and tuning in the cochlea. Neuroscientist. 2003;9:311–316. - PubMed
-
- Mann ZF, Kelley MW. Development of tonotopy in the auditory periphery. Hear Res. 2011;276:2–15. - PubMed
-
- Meyer AC, Frank T, Khimich D, Hoch G, Riedel D, et al. Tuning of synapse number, structure and function in the cochlea. Nat Neurosci. 2009;12:444–453. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
