

Campylobacter jejuni induces acute enterocolitis in gnotobiotic IL-10-/- mice via Toll-like-receptor-2 and -4 signaling
- PMID: 22808254
- PMCID: PMC3393706
- DOI: 10.1371/journal.pone.0040761
Campylobacter jejuni induces acute enterocolitis in gnotobiotic IL-10-/- mice via Toll-like-receptor-2 and -4 signaling
Abstract
Background: Campylobacter jejuni is a leading cause of foodborne bacterial enterocolitis worldwide. Investigation of immunopathology is hampered by a lack of suitable vertebrate models. We have recently shown that gnotobiotic mice as well as conventional IL-10(-/-) animals are susceptible to C. jejuni infection and develop intestinal immune responses. However, clinical symptoms of C. jejuni infection were rather subtle and did not reflect acute bloody diarrhea seen in human campylobacteriosis.
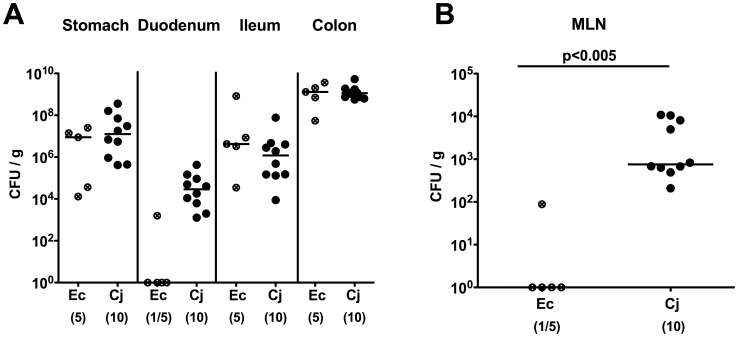
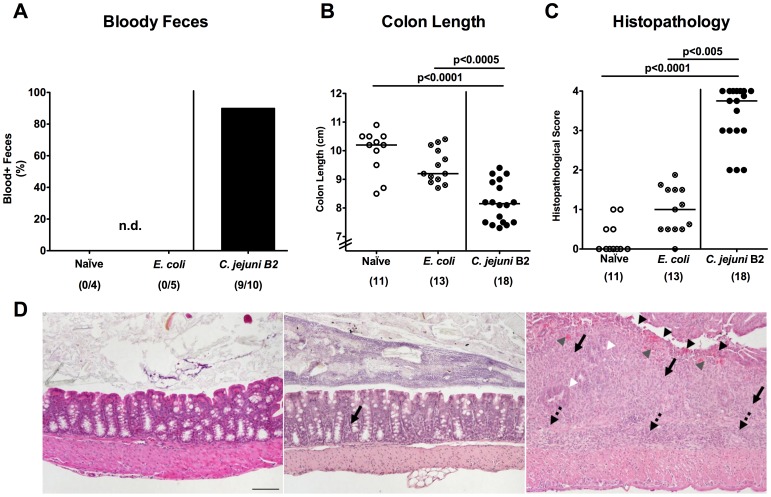
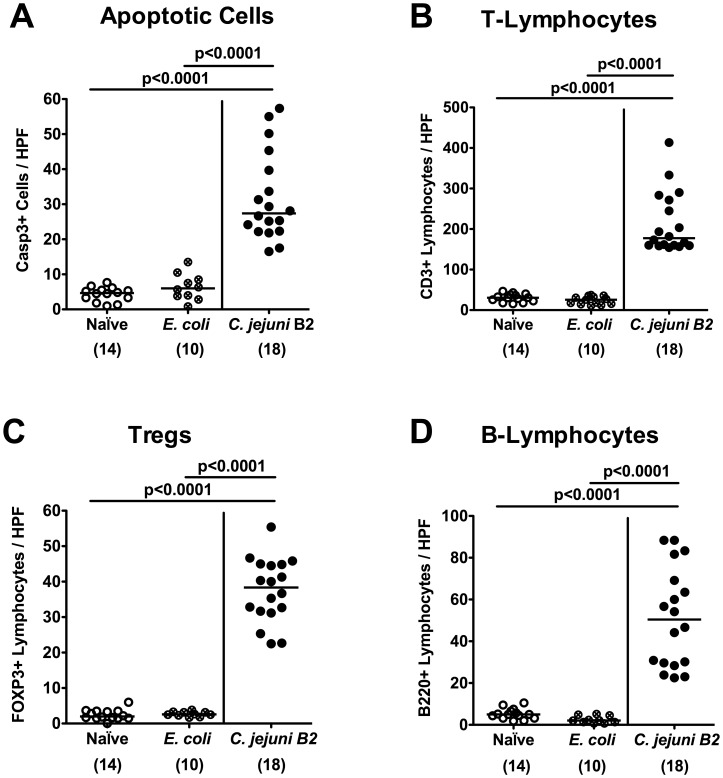
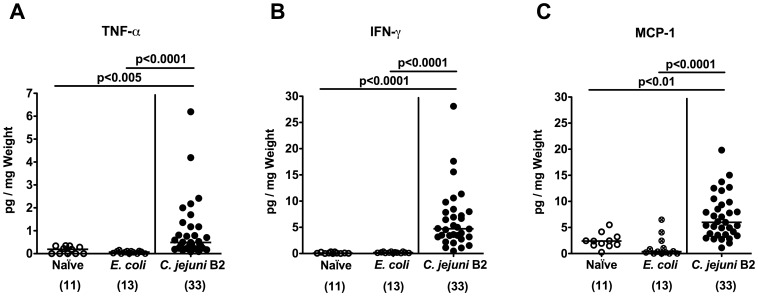
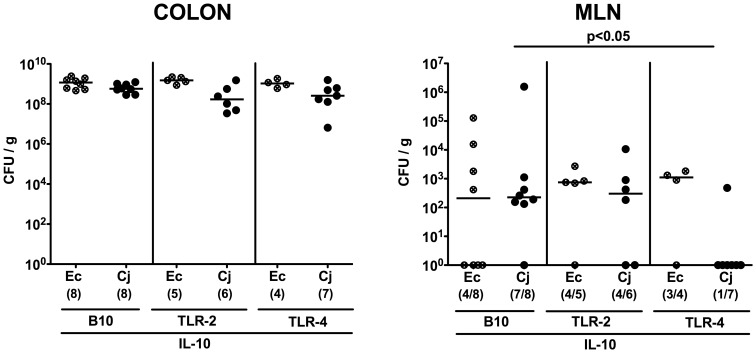
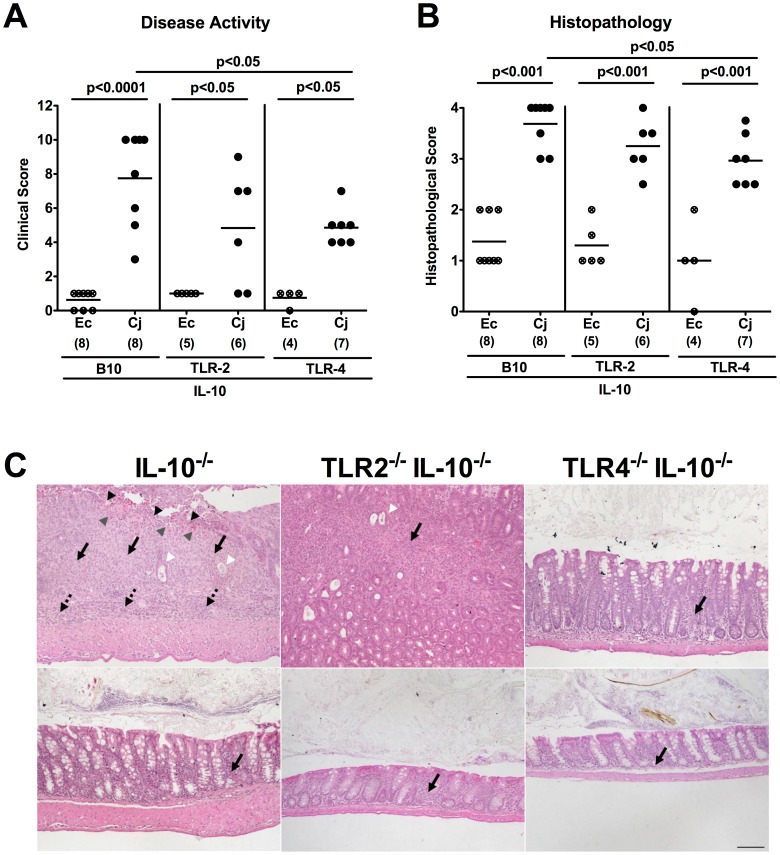
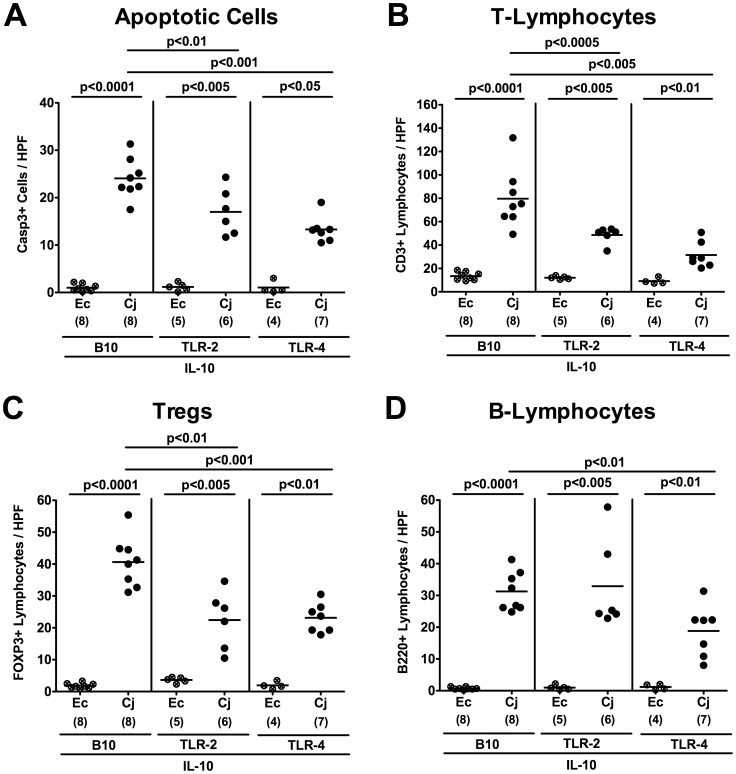
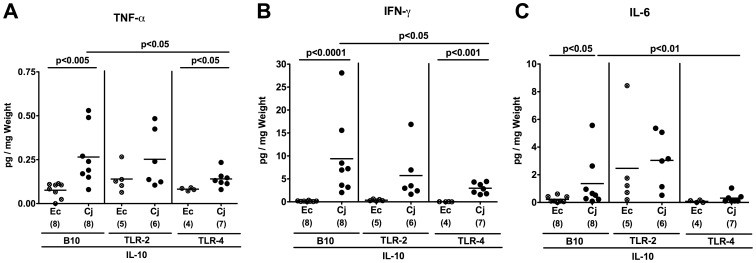
Methodology/principal findings: In order to overcome these limitations we generated gnotobiotic IL-10(-/-) mice by quintuple antibiotic treatment starting right after weaning. The early treatment was essential to prevent these animals from chronic colitis. Following oral infection C. jejuni colonized the gastrointestinal tract at high levels and induced acute enterocolitis within 7 days as indicated by bloody diarrhea and pronounced histopathological changes of the colonic mucosa. Immunopathology was further characterized by increased numbers of apoptotic cells, regulatory T-cells, T- and B-lymphocytes as well as elevated TNF-α, IFN-γ, and MCP-1 concentrations in the inflamed colon. The induction of enterocolitis was specific for C. jejuni given that control animals infected with a commensal E. coli strain did not display any signs of disease. Most strikingly, intestinal immunopathology was ameliorated in mice lacking Toll-like-receptors-2 or -4 indicating that C. jejuni lipoproteins and lipooligosaccharide are essential for induction and progression of immunopathology.
Conclusion/significance: Gnotobiotic IL-10(-/-) mice develop acute enterocolitis following C. jejuni infection mimicking severe episodes of human campylobacteriosis and are thus well suited to further dissect mechanisms underlying Campylobacter infections in vivo.
Conflict of interest statement
Figures
References
-
- Hermans D, Pasmans F, Messens W, Martel A, Van Immerseel F, et al. Poultry as a Host for the Zoonotic Pathogen Campylobacter jejuni. Vector Borne Zoonotic Dis. 2012. - PubMed
-
- Lane JA, Mehra RK, Carrington SD, Hickey RM. The food glycome: a source of protection against pathogen colonization in the gastrointestinal tract. Int J Food Microbiol. 2010;142:1–13. - PubMed
-
- Young KT, Davis LM, Dirita VJ. Campylobacter jejuni: molecular biology and pathogenesis. Nat Rev Microbiol. 2007;5:665–679. - PubMed
-
- Havelaar AH, van Pelt W, Ang CW, Wagenaar JA, van Putten JP, et al. Immunity to Campylobacter: its role in risk assessment and epidemiology. Crit Rev Microbiol. 2009;35:1–22. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
