

Role of the gut in lipid homeostasis
- PMID: 22811425
- PMCID: PMC3589762
- DOI: 10.1152/physrev.00019.2011
Role of the gut in lipid homeostasis
Abstract
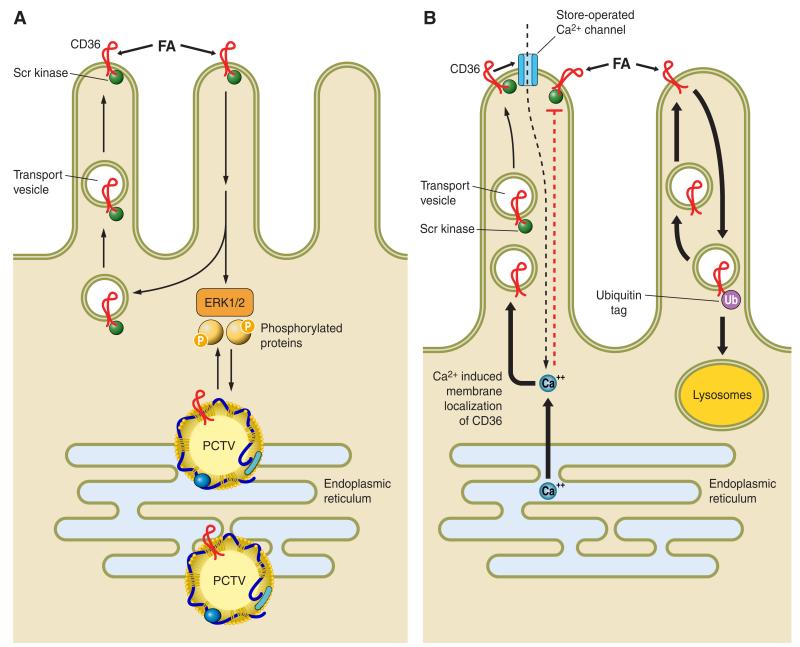
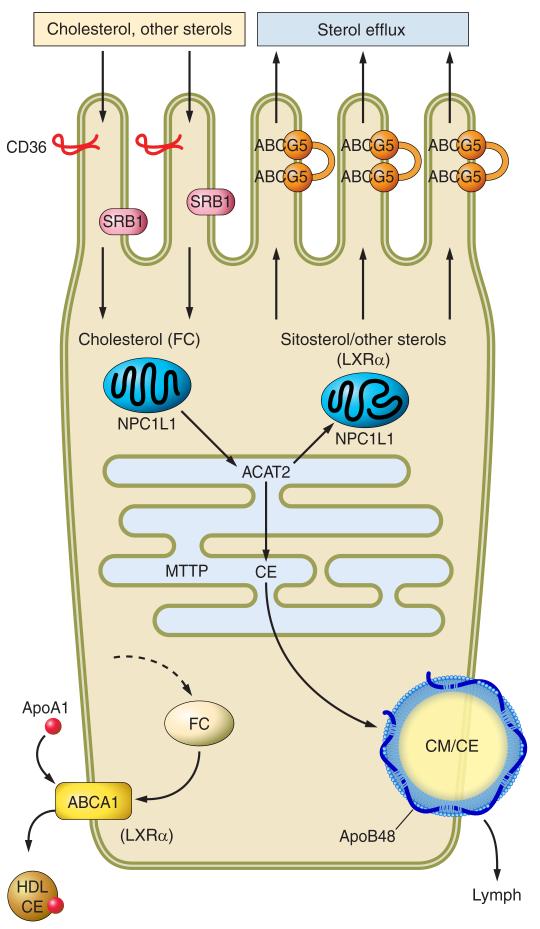
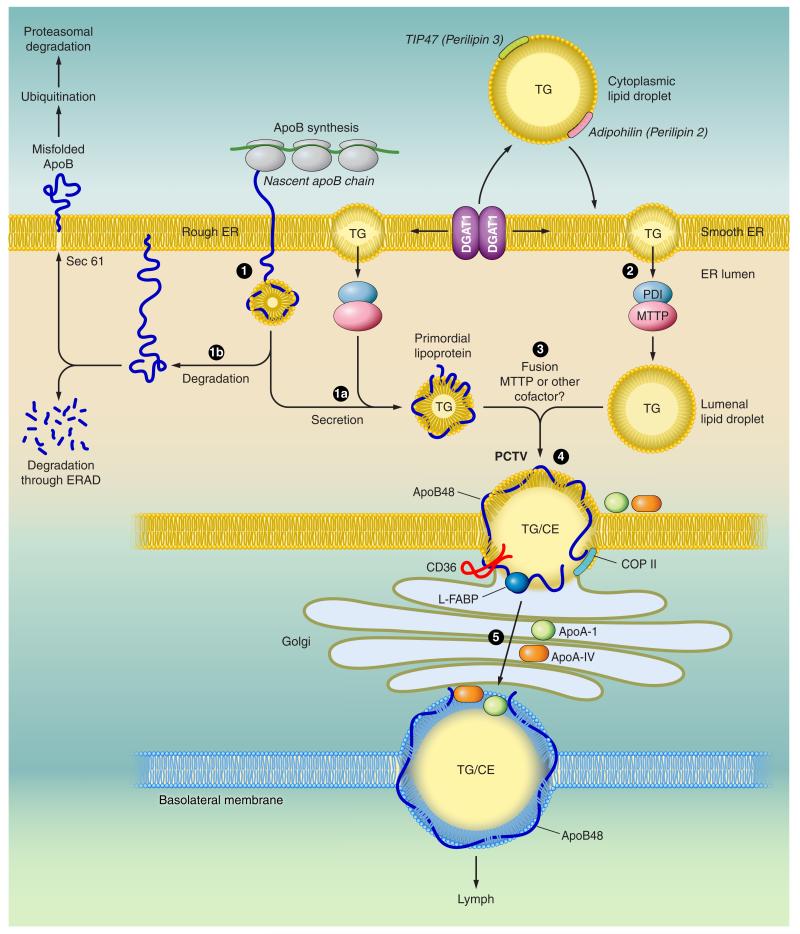
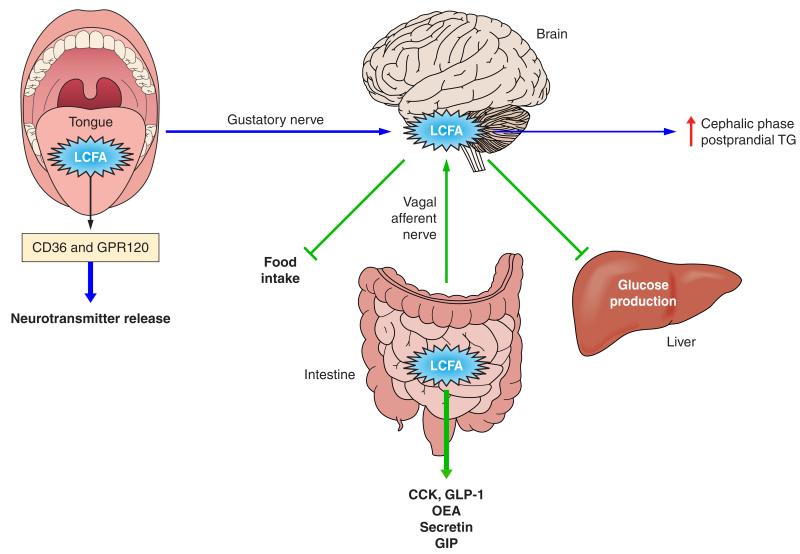
Intestinal lipid transport plays a central role in fat homeostasis. Here we review the pathways regulating intestinal absorption and delivery of dietary and biliary lipid substrates, principally long-chain fatty acid, cholesterol, and other sterols. We discuss the regulation and functions of CD36 in fatty acid absorption, NPC1L1 in cholesterol absorption, as well as other lipid transporters including FATP4 and SRB1. We discuss the pathways of intestinal sterol efflux via ABCG5/G8 and ABCA1 as well as the role of the small intestine in high-density lipoprotein (HDL) biogenesis and reverse cholesterol transport. We review the pathways and genetic regulation of chylomicron assembly, the role of dominant restriction points such as microsomal triglyceride transfer protein and apolipoprotein B, and the role of CD36, l-FABP, and other proteins in formation of the prechylomicron complex. We will summarize current concepts of regulated lipoprotein secretion (including HDL and chylomicron pathways) and include lessons learned from families with genetic mutations in dominant pathways (i.e., abetalipoproteinemia, chylomicron retention disease, and familial hypobetalipoproteinemia). Finally, we will provide an integrative view of intestinal lipid homeostasis through recent findings on the role of lipid flux and fatty acid signaling via diverse receptor pathways in regulating absorption and production of satiety factors.
Figures
References
-
- Abumrad NA, el-Maghrabi MR, Amri EZ, Lopez E, Grimaldi PA. Cloning of a rat adipocyte membrane protein implicated in binding or transport of long-chain fatty acids that is induced during preadipocyte differentiation. Homology with human CD36. J Biol Chem. 1993;268:17665–17668. - PubMed
-
- Ailhaud G, Guesnet P, Cunnane SC. An emerging risk factor for obesity: does disequilibrium of polyunsaturated fatty acid metabolism contribute to excessive adipose tissue development? Br J Nutr. 2008;100:461–470. - PubMed
-
- Akesson B, Gronowitz S, Herslof B, Ohlson R. Absorption of synthetic, stereochemically defined acylglycerols in the rat. Lipids. 1978;13:338–343. - PubMed
-
- Al Massadi O, Tschop MH, Tong J. Ghrelin acylation and metabolic control. Peptides. 2011;32:2301–2308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK060022/DK/NIDDK NIH HHS/United States
- HL-057278C/HL/NHLBI NIH HHS/United States
- R01 HL038180/HL/NHLBI NIH HHS/United States
- HL-38180/HL/NHLBI NIH HHS/United States
- R37 HL038180/HL/NHLBI NIH HHS/United States
- DK-60022/DK/NIDDK NIH HHS/United States
- DK-52574/DK/NIDDK NIH HHS/United States
- R01 DK033301/DK/NIDDK NIH HHS/United States
- DK-56260/DK/NIDDK NIH HHS/United States
- DK-033301/DK/NIDDK NIH HHS/United States
- P01 HL057278/HL/NHLBI NIH HHS/United States
- R01 DK056260/DK/NIDDK NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical