

Three murine leukemia virus integration regions within 100 kilobases upstream of c-myb are proximal to the 5' regulatory region of the gene through DNA looping
- PMID: 22811527
- PMCID: PMC3457256
- DOI: 10.1128/JVI.01077-12
Three murine leukemia virus integration regions within 100 kilobases upstream of c-myb are proximal to the 5' regulatory region of the gene through DNA looping
Abstract
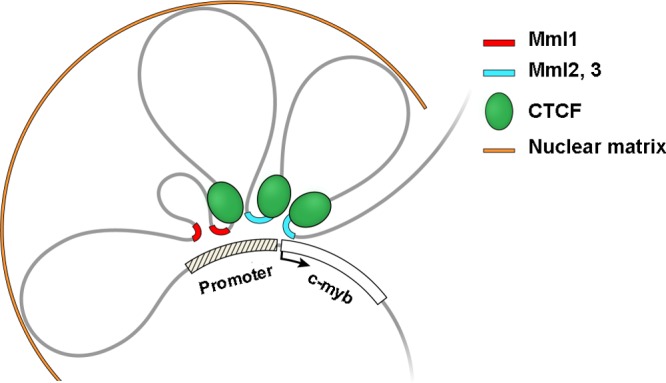
Retroviruses integrated into genomic DNA participate in long-range gene activation from as far away as several hundred kilobases. Hypotheses have been put forth to account for these phenomena, but data have not been provided to support a physical mechanism that explains long-range activation. In murine leukemia virus-induced myeloid leukemia in mice, integrated proviruses have been found upstream of c-myb in three regions, named Mml1, Mml2, and Mml3 (25, 50, and 70 kb upstream, respectively). The transcription factor c-Myb is an oncogene whose dysregulation and/or mutation can lead to human leukemia. We hypothesized that the murine c-myb upstream region contains regulatory elements accessed by the retrovirus. To identify regulatory sites in the murine c-myb upstream region, we looked by chromatin immunoprecipitation with microarray technology (ChIP-on-chip) for histone modifications implicating gene activation in normal cells. H3K4me3, H3K4me1, and H3K9/14ac were enriched at Mml1 and/or Mml2 in the myeloblastic cell line M1, which expresses c-myb. The enrichment of all of these histone marks decreased with differentiation-induced downregulation of the gene in M1 cells but increased and spread in tumor cells containing integrated provirus. Importantly, using chromosome conformation capture (3C)-quantitative PCR assays, interactions between the 5' region, including the promoter and all Mml sites (Mml1, Mml2, and Mml3), were detected due to DNA looping in M1 cells and tumor cells with provirus in Mml1, Mml2, or Mml3. Therefore, our study provides a new mechanism of retrovirus insertional mutagenesis whereby spatial chromatin organization allows distally located provirus, with its own enhancer elements, to access the 5' regulatory region of the gene.
Figures





References
-
- Barski A, et al. 2007. High-resolution profiling of histone methylations in the human genome. Cell 129:823–837 - PubMed
-
- Bender TP, Thompson CB, Kuehl WM. 1987. Differential expression of c-myb mRNA in murine B lymphomas by a block to transcription elongation. Science 237:1473–1476 - PubMed
-
- Clappier E, et al. 2007. The C-MYB locus is involved in chromosomal translocation and genomic duplications in human T-cell acute leukemia (T-ALL), the translocation defining a new T-ALL subtype in very young children. Blood 110:1251–1261 - PubMed
-
- Dekker J, Rippe K, Dekker M, Kleckner N. 2002. Capturing chromosome conformation. Science 295:1306–1311 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
