

Glutaredoxin 1 protects dopaminergic cells by increased protein glutathionylation in experimental Parkinson's disease
- PMID: 22816731
- PMCID: PMC3474191
- DOI: 10.1089/ars.2011.4474
Glutaredoxin 1 protects dopaminergic cells by increased protein glutathionylation in experimental Parkinson's disease
Abstract
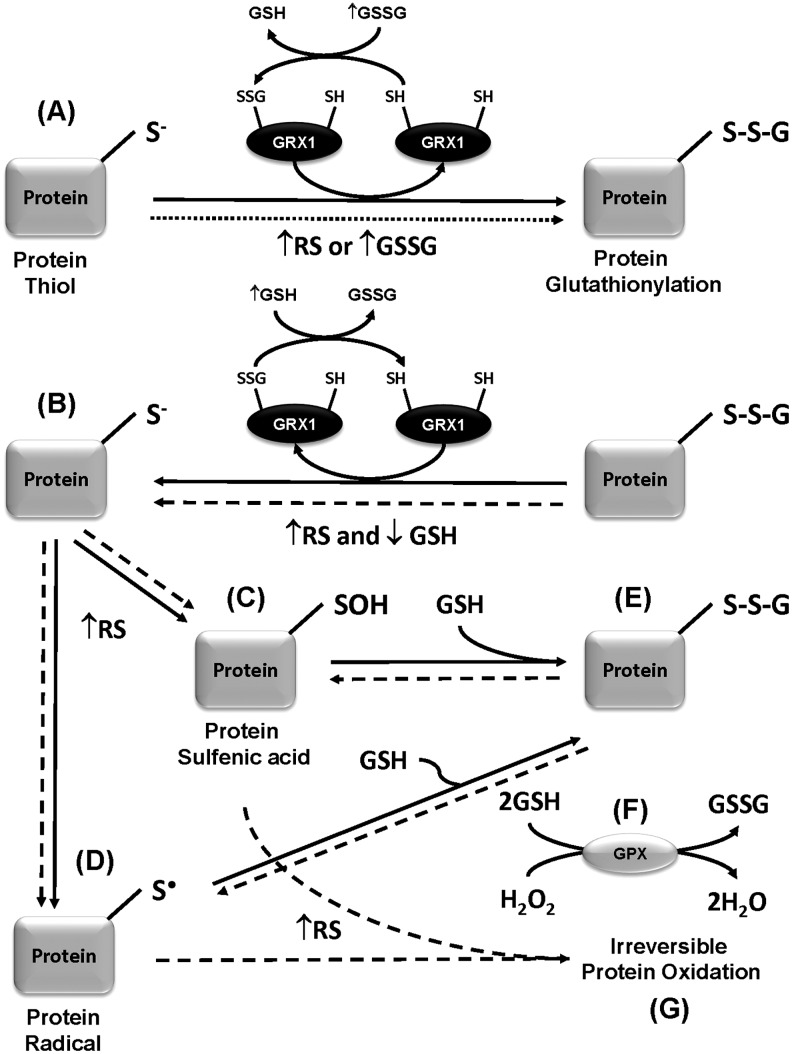
Aims: Chronic exposure to environmental toxicants, such as paraquat, has been suggested as a risk factor for Parkinson's disease (PD). Although dopaminergic cell death in PD is associated with oxidative damage, the molecular mechanisms involved remain elusive. Glutaredoxins (GRXs) utilize the reducing power of glutathione to modulate redox-dependent signaling pathways by protein glutathionylation. We aimed to determine the role of GRX1 and protein glutathionylation in dopaminergic cell death.
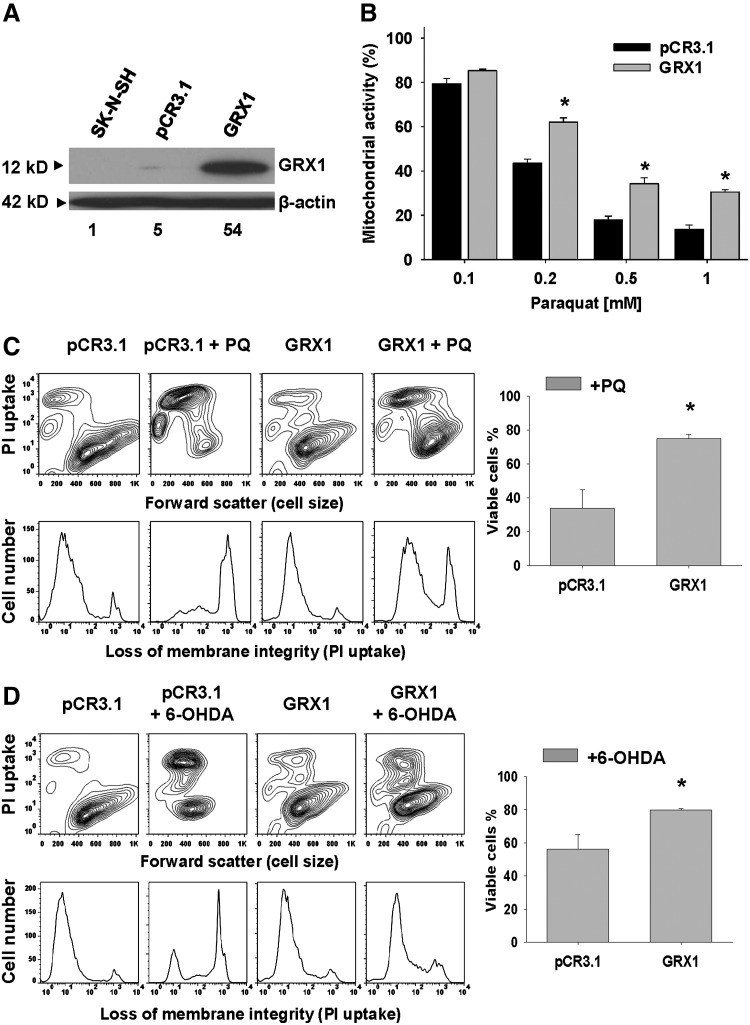
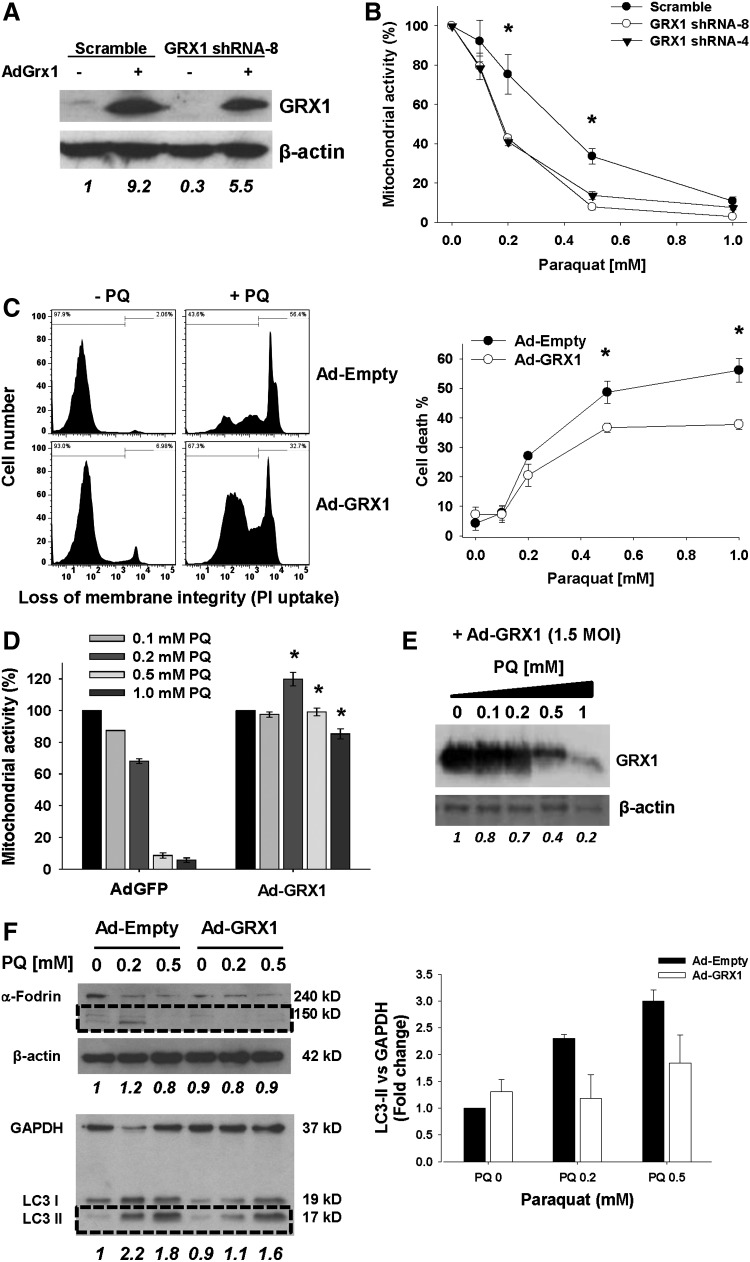
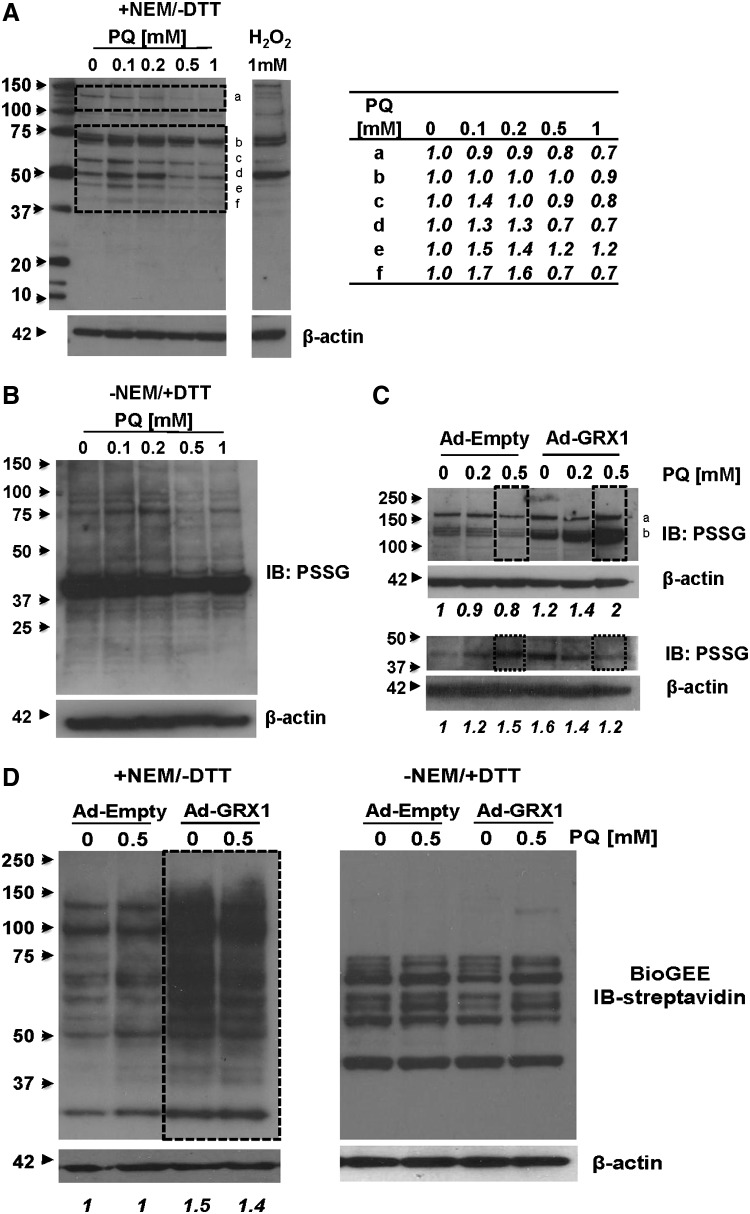
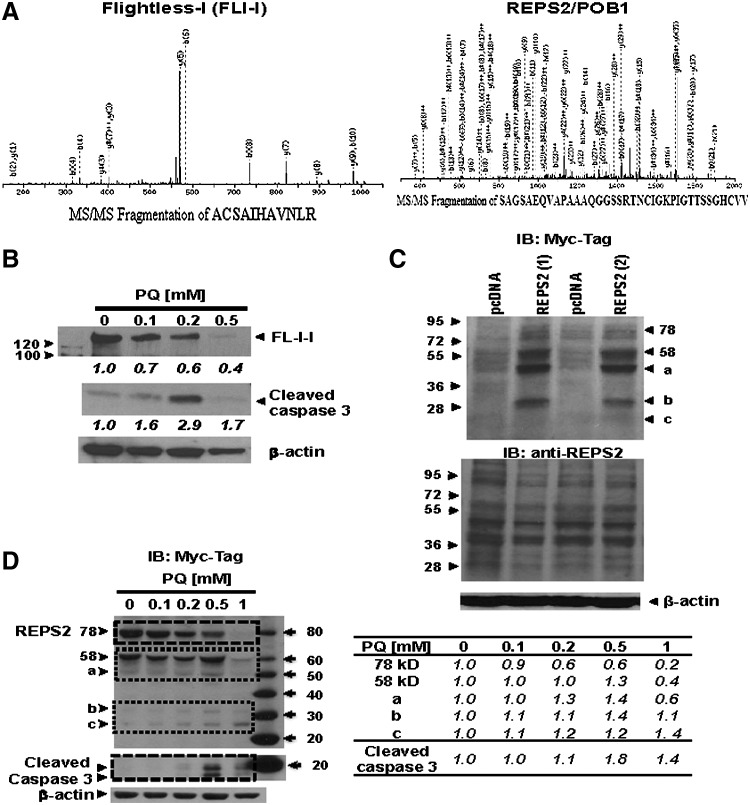
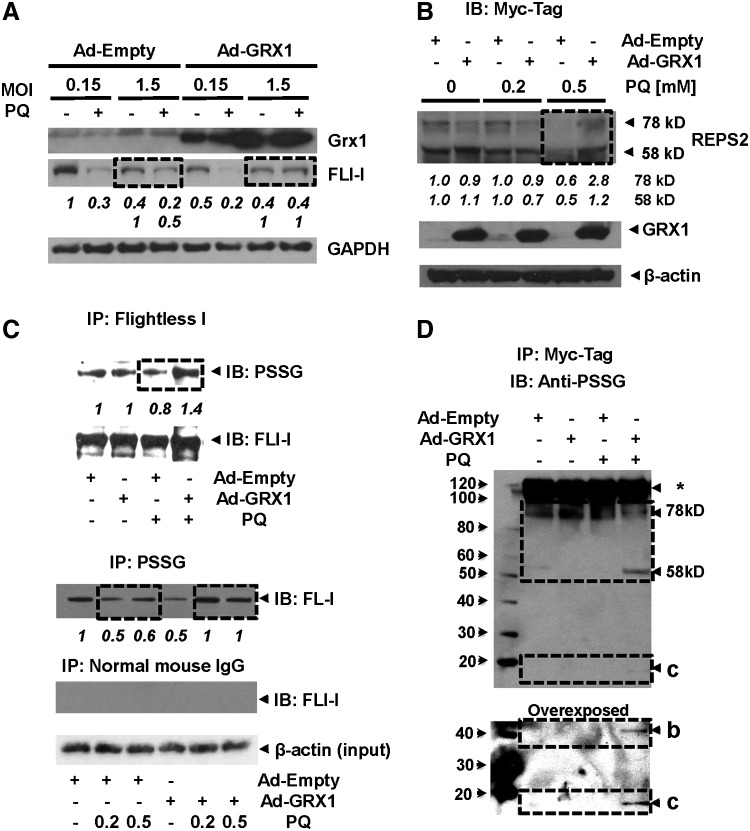
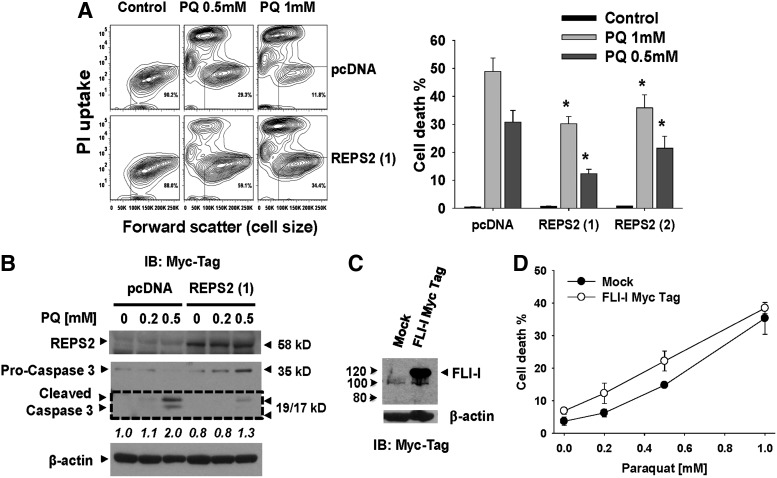
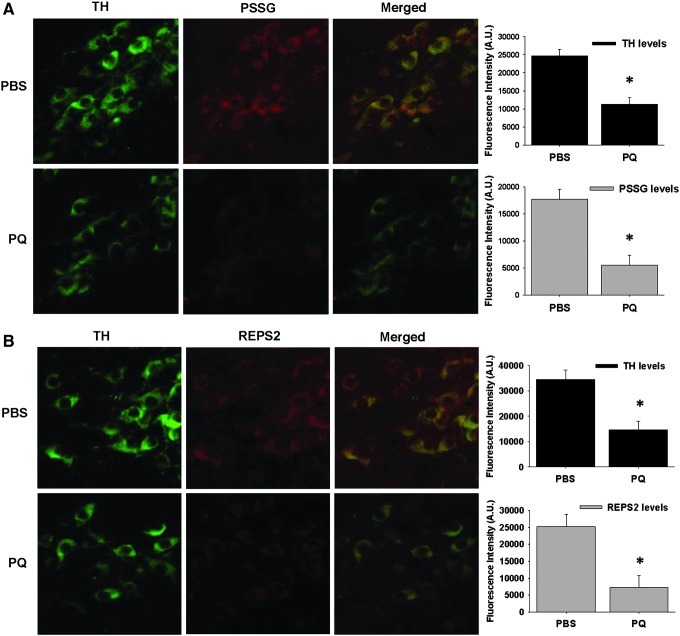
Results: In dopaminergic cells, toxicity induced by paraquat or 6-hydroxydopamine (6-OHDA) was inhibited by GRX1 overexpression, while its knock-down sensitized cells to paraquat-induced cell death. Dopaminergic cell death was paralleled by protein deglutathionylation, and this was reversed by GRX1. Mass spectrometry analysis of immunoprecipitated glutathionylated proteins identified the actin binding flightless-1 homolog protein (FLI-I) and the RalBP1-associated Eps domain-containing protein 2 (REPS2/POB1) as targets of glutathionylation in dopaminergic cells. Paraquat induced the degradation of FLI-I and REPS2 proteins, which corresponded with the activation of caspase 3 and cell death progression. GRX1 overexpression reduced both the degradation and deglutathionylation of FLI-I and REPS2, while stable overexpression of REPS2 reduced paraquat toxicity. A decrease in glutathionylated proteins and REPS2 levels was also observed in the substantia nigra of mice treated with paraquat.
Innovation: We have identified novel protein targets of glutathionylation in dopaminergic cells and demonstrated the protective role of GRX1-mediated protein glutathionylation against paraquat-induced toxicity.
Conclusions: These results demonstrate a protective role for GRX1 and increased protein glutathionylation in dopaminergic cell death induced by paraquat, and identify a novel protective role for REPS2.
Figures
References
-
- Alam ZI. Daniel SE. Lees AJ. Marsden DC. Jenner P. Halliwell B. A generalised increase in protein carbonyls in the brain in Parkinson's but not incidental Lewy body disease. J Neurochem. 1997;69:1326–1329. - PubMed
-
- Alam ZI. Jenner A. Daniel SE. Lees AJ. Cairns N. Marsden CD. Jenner P. Halliwell B. Oxidative DNA damage in the parkinsonian brain: An apparent selective increase in 8-hydroxyguanine levels in substantia nigra. J Neurochem. 1997;69:1196–1203. - PubMed
-
- Aon-Bertolino ML. Romero JI. Galeano P. Holubiec M. Badorrey MS. Saraceno GE. Hanschmann EM. Lillig CH. Capani F. Thioredoxin and glutaredoxin system proteins-immunolocalization in the rat central nervous system. Biochim Biophys Acta. 2011;1810:93–110. - PubMed
-
- Archer SK. Claudianos C. Campbell HD. Evolution of the gelsolin family of actin-binding proteins as novel transcriptional coactivators. Bioessays. 2005;27:388–396. - PubMed
-
- Ascherio A. Chen H. Weisskopf MG. O'Reilly E. McCullough ML. Calle EE. Schwarzschild MA. Thun MJ. Pesticide exposure and risk for Parkinson's disease. Ann Neurol. 2006;60:197–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
