

Memory CD4+ T cells protect against influenza through multiple synergizing mechanisms
- PMID: 22820287
- PMCID: PMC3408751
- DOI: 10.1172/JCI63689
Memory CD4+ T cells protect against influenza through multiple synergizing mechanisms
Abstract
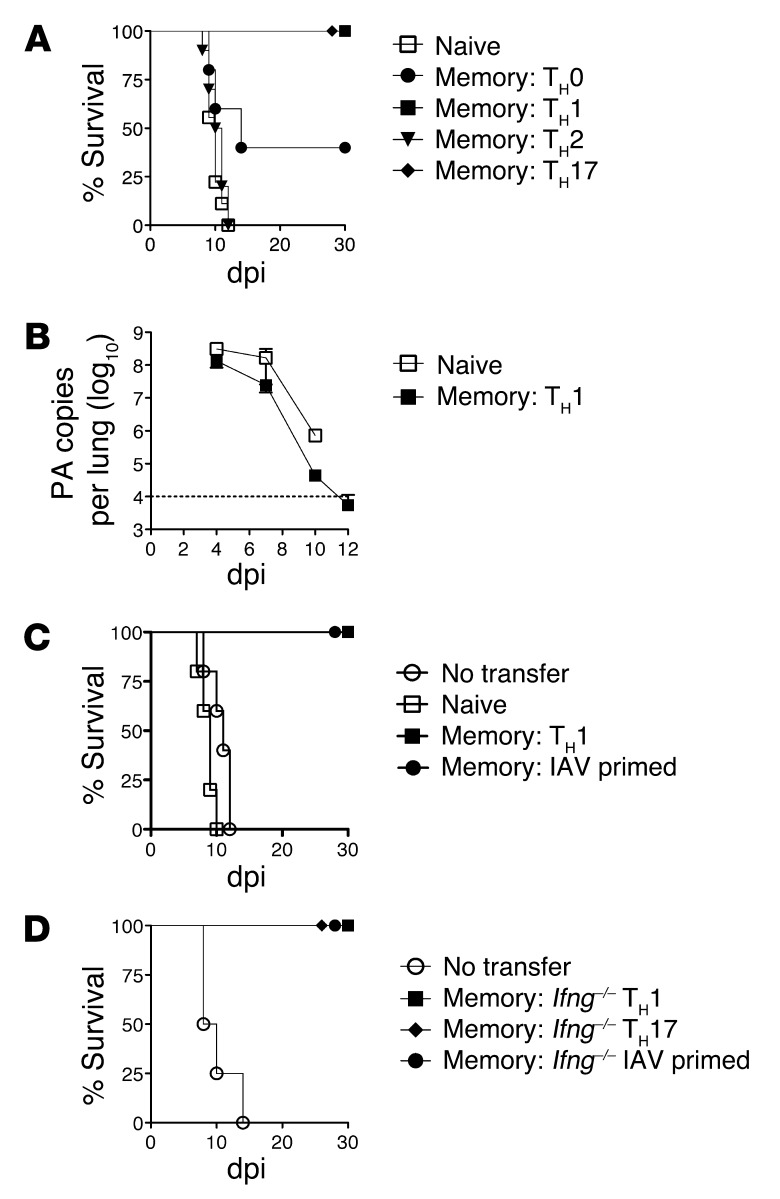
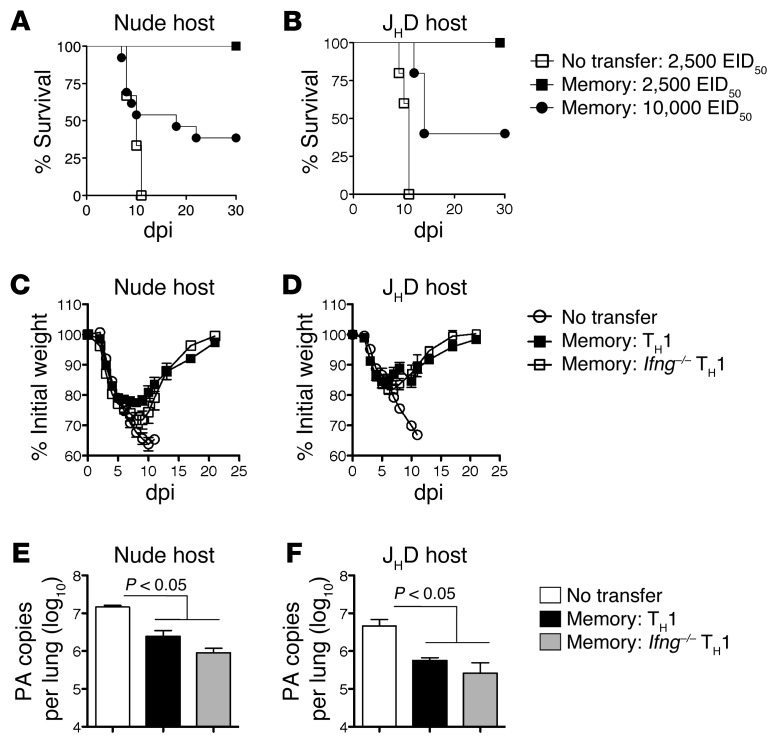
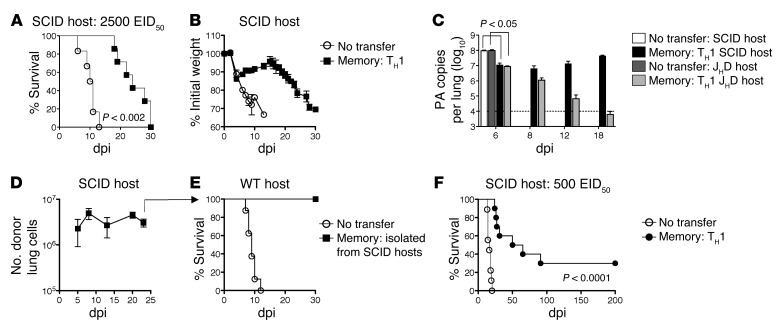
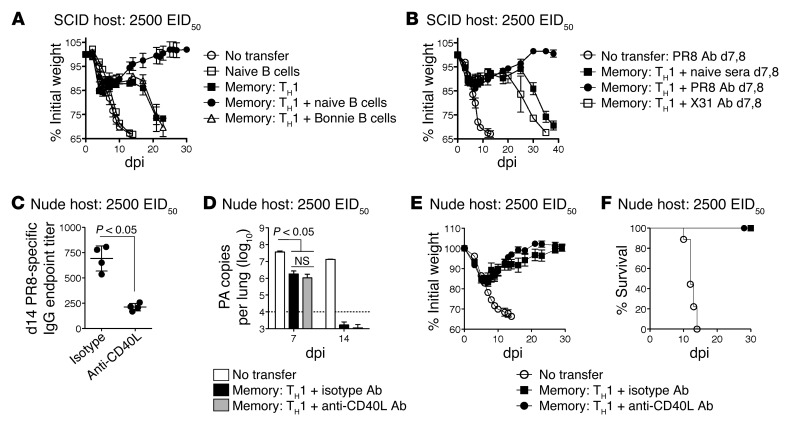
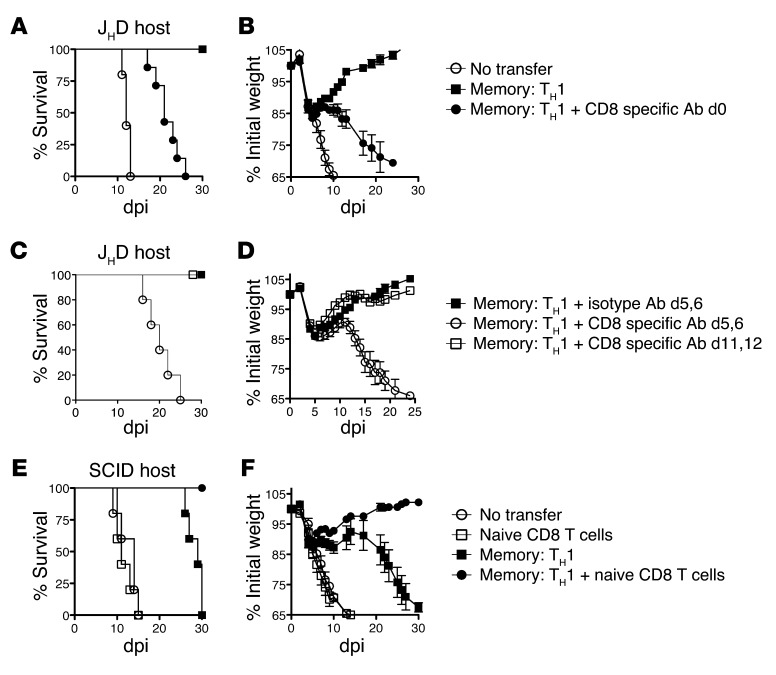
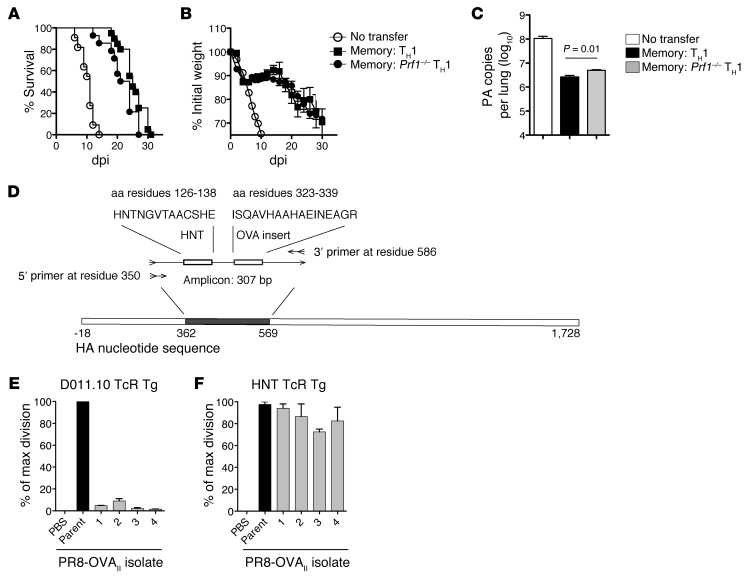
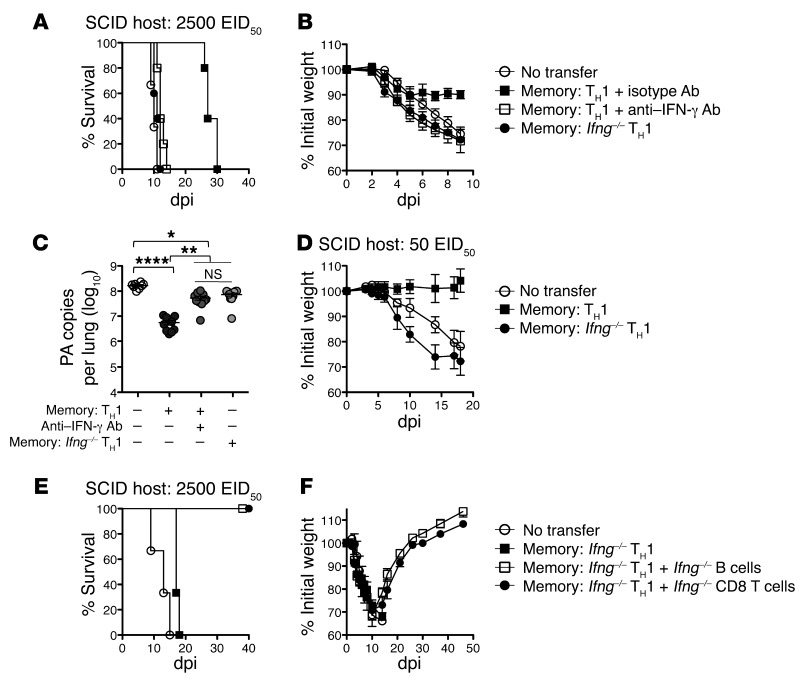
Memory CD4+ T cells combat viral infection and contribute to protective immune responses through multiple mechanisms, but how these pathways interact is unclear. We found that several pathways involving memory CD4+ T cells act together to effectively clear influenza A virus (IAV) in otherwise unprimed mice. Memory CD4+ T cell protection was enhanced through synergy with naive B cells or CD8+ T cells and maximized when both were present. However, memory CD4+ T cells protected against lower viral doses independently of other lymphocytes through production of IFN-γ. Moreover, memory CD4+ T cells selected for epitope-specific viral escape mutants via a perforin-dependent pathway. By deconstructing protective immunity mediated by memory CD4+ T cells, we demonstrated that this population simultaneously acts through multiple pathways to provide a high level of protection that ensures eradication of rapidly mutating pathogens such as IAV. This redundancy indicates the need for reductionist approaches for delineating the individual mechanisms of protection mediated by memory CD4+ T cells responding to pathogens.
Figures
Comment in
-
Memory CD4+ T cells: beyond "helper" functions.J Clin Invest. 2012 Aug;122(8):2768-70. doi: 10.1172/JCI65208. Epub 2012 Jul 23. J Clin Invest. 2012. PMID: 22820285 Free PMC article.
-
Antiviral immunity: Deconstructing CD4+ T cell memory.Nat Rev Immunol. 2012 Sep;12(9):618-9. doi: 10.1038/nri3290. Nat Rev Immunol. 2012. PMID: 22918460 No abstract available.
References
-
- Allan W, Tabi Z, Cleary A, Doherty PC. Cellular events in the lymph node and lung of mice with influenza. Consequences of depleting CD4+ T cells. J Immunol. 1990;144(10):3980–3986. - PubMed
-
- Brown DM, Dilzer AM, Meents DL, Swain SL. CD4 T cell-mediated protection from lethal influenza: perforin and antibody-mediated mechanisms give a one-two punch. J Immunol. 2006;177(5):2888–2898. - PubMed
-
- Powell TJ, et al. Priming with cold-adapted influenza A does not prevent infection but elicits long-lived protection against supralethal challenge with heterosubtypic virus. J Immunol. 2007;178(2):1030–1038. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
