

Direct sequencing of Arabidopsis thaliana RNA reveals patterns of cleavage and polyadenylation
- PMID: 22820990
- PMCID: PMC3533403
- DOI: 10.1038/nsmb.2345
Direct sequencing of Arabidopsis thaliana RNA reveals patterns of cleavage and polyadenylation
Abstract
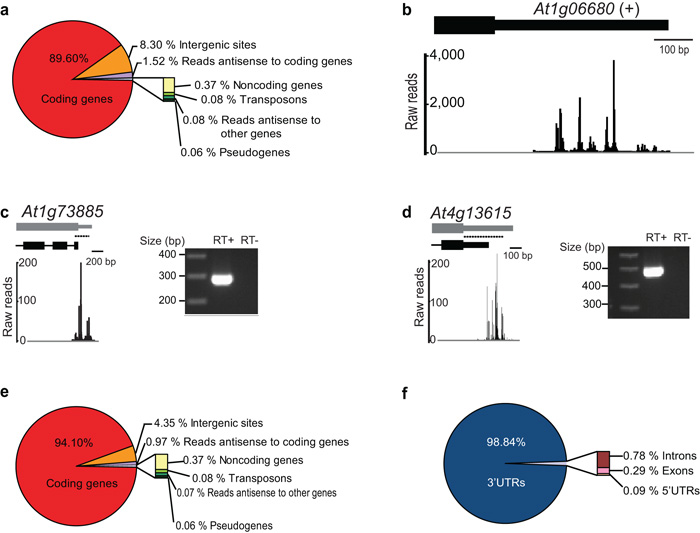
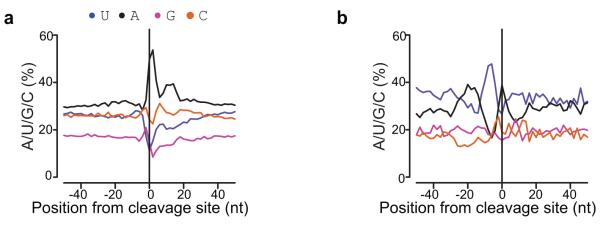
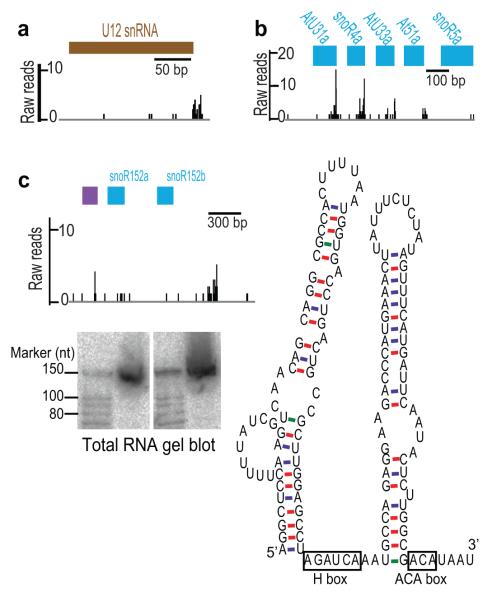
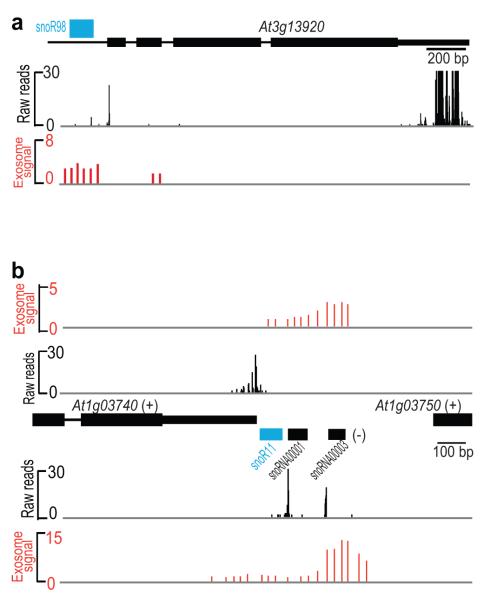
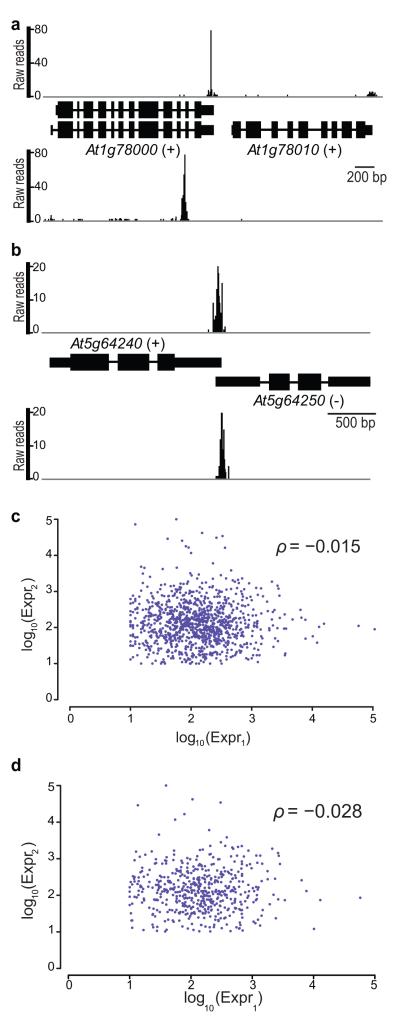
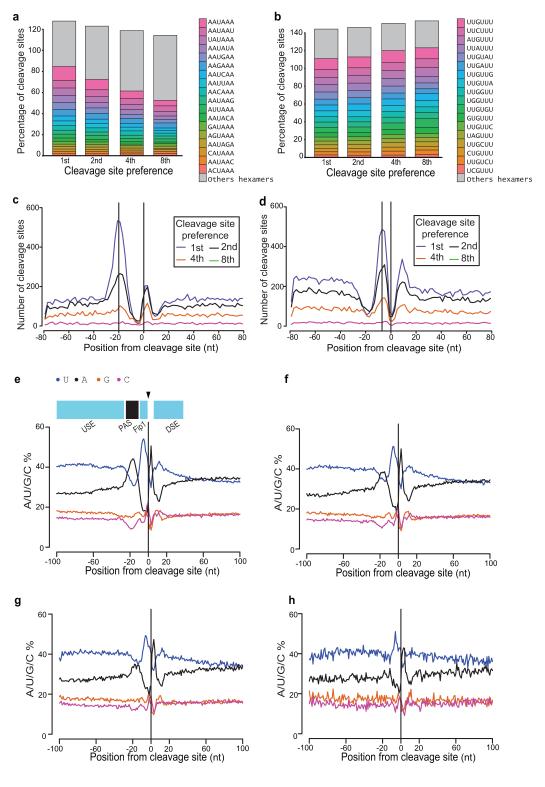
It has recently been shown that RNA 3'-end formation plays a more widespread role in controlling gene expression than previously thought. To examine the impact of regulated 3'-end formation genome-wide, we applied direct RNA sequencing to A. thaliana. Here we show the authentic transcriptome in unprecedented detail and describe the effects of 3'-end formation on genome organization. We reveal extreme heterogeneity in RNA 3' ends, discover previously unrecognized noncoding RNAs and propose widespread reannotation of the genome. We explain the origin of most poly(A)(+) antisense RNAs and identify cis elements that control 3'-end formation in different registers. These findings are essential to understanding what the genome actually encodes, how it is organized and how regulated 3'-end formation affects these processes.
Figures

References
-
- Hornyik C, Terzi LC, Simpson GG. The spen family protein FPA controls alternative cleavage and polyadenylation of RNA. Dev. Cell. 2010;18:203–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
