

Comparative interaction networks: bridging genotype to phenotype
- PMID: 22821457
- PMCID: PMC3518490
- DOI: 10.1007/978-1-4614-3567-9_7
Comparative interaction networks: bridging genotype to phenotype
Abstract
Over the past decade, biomedical research has witnessed an exponential increase in the throughput of the characterization of biological systems. Here we review the recent progress in large-scale methods to determine protein-protein, genetic and chemical-genetic interaction networks. We discuss some of the limitations and advantages of the different methods and give examples of how these networks are being used to study the evolutionary process. Comparative studies have revealed that different types of protein-protein interactions diverge at different rates with high conservation of co-complex membership but rapid divergence of more promiscuous interactions like those that mediate post-translational modifications. These evolutionary trends have consistent genetic consequences with highly conserved epistatic interactions within complex subunits but faster divergence of epistatic interactions across complexes or pathways. Finally, we discuss how these evolutionary observations are being used to interpret cross-species chemical-genetic studies and how they might shape therapeutic strategies. Together, these interaction networks offer us an unprecedented level of detail into how genotypes are translated to phenotypes, and we envision that they will be increasingly useful in the interpretation of genetic and phenotypic variation occurring within populations as well as the rational design of combinatorial therapeutics.
Figures
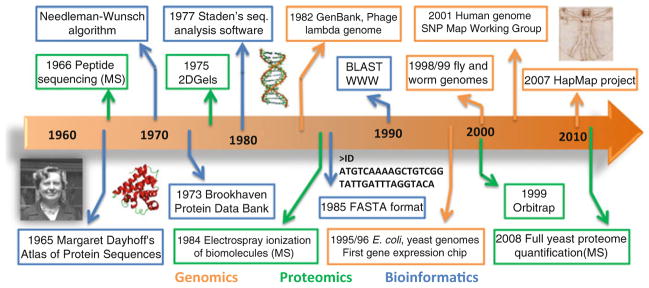
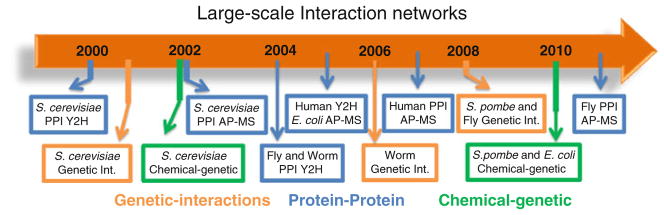
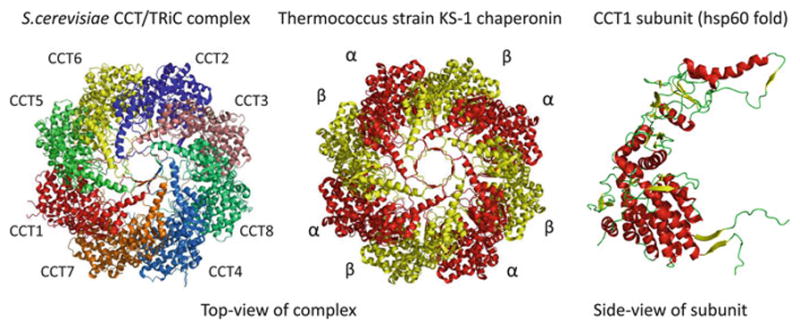
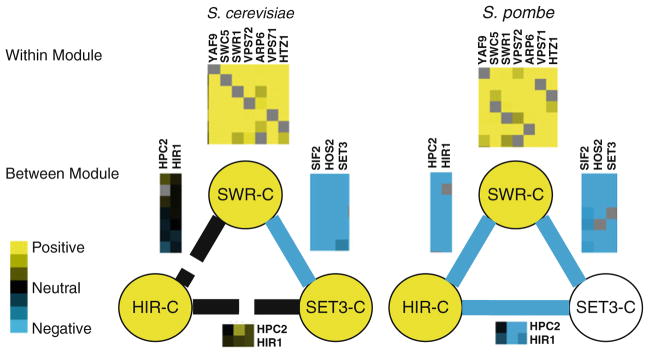
References
-
- Lynch M. The evolutionary fate and consequences of duplicate genes. Science. 2000;290:1151–1155. - PubMed
-
- Dietrich FS, Voegeli S, Brachat S, Lerch A, Gates K, et al. The Ashbya gossypii genome as a tool for mapping the ancient Saccharomyces cerevisiae genome. Science (New York, NY) 2004;304:304–307. - PubMed
-
- Kellis M, Birren BW, Lander ES. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae. Nature. 2004;428:617–624. - PubMed
-
- Lynch M, Conery JS. The origins of genome complexity. Science (New York, NY) 2003;302:1401–1404. - PubMed
Publication types
MeSH terms
Grants and funding
- GM082250/GM/NIGMS NIH HHS/United States
- GM084279/GM/NIGMS NIH HHS/United States
- P50 GM081879/GM/NIGMS NIH HHS/United States
- GM084448/GM/NIGMS NIH HHS/United States
- R01 GM084279/GM/NIGMS NIH HHS/United States
- P01 AI091575/AI/NIAID NIH HHS/United States
- AI090935/AI/NIAID NIH HHS/United States
- R01 GM098101/GM/NIGMS NIH HHS/United States
- GM098101/GM/NIGMS NIH HHS/United States
- P50 GM082250/GM/NIGMS NIH HHS/United States
- P30 AI027763/AI/NIAID NIH HHS/United States
- R01 GM084448/GM/NIGMS NIH HHS/United States
- R01 GM078360/GM/NIGMS NIH HHS/United States
- P01 AI090935/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
