

Scavenger receptor A restrains T-cell activation and protects against concanavalin A-induced hepatic injury
- PMID: 22821642
- PMCID: PMC3491179
- DOI: 10.1002/hep.25983
Scavenger receptor A restrains T-cell activation and protects against concanavalin A-induced hepatic injury
Abstract
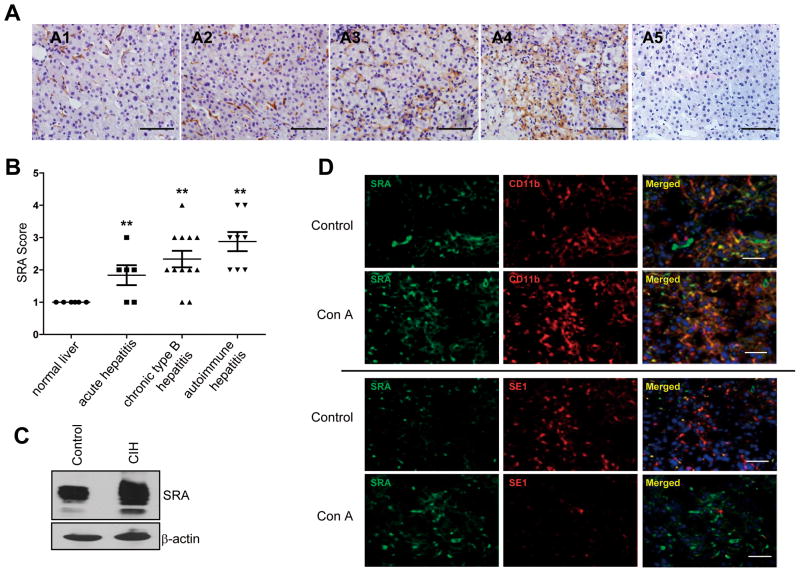
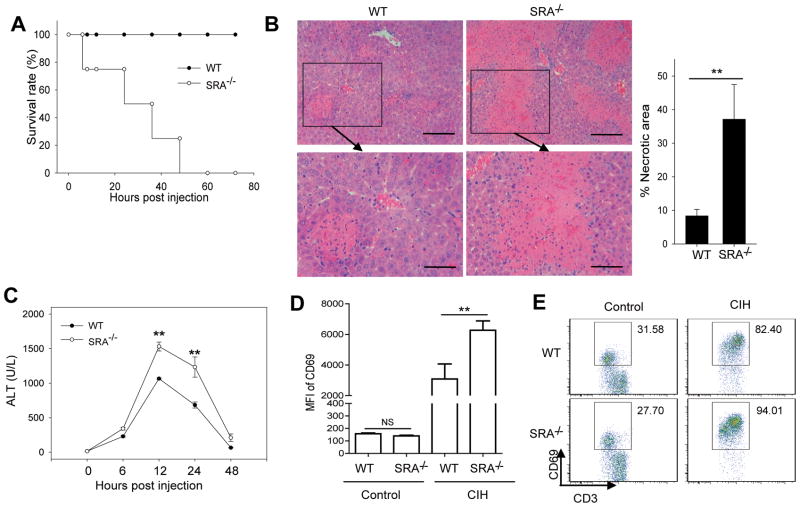
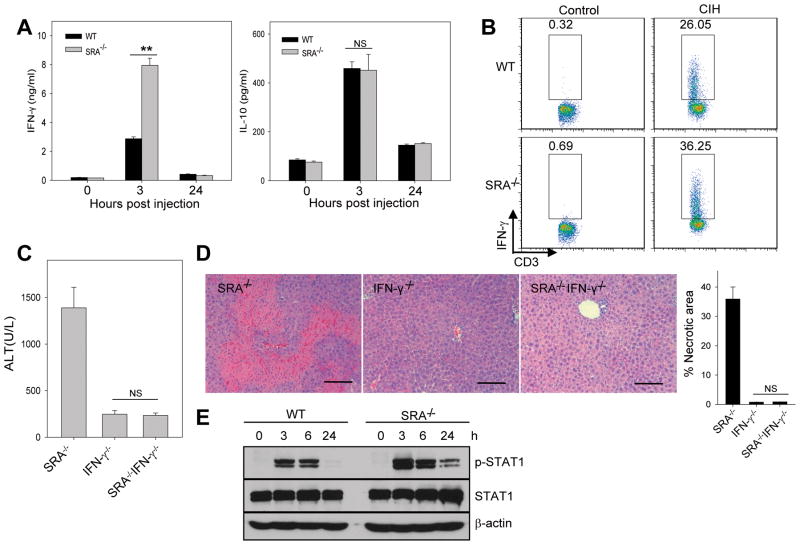
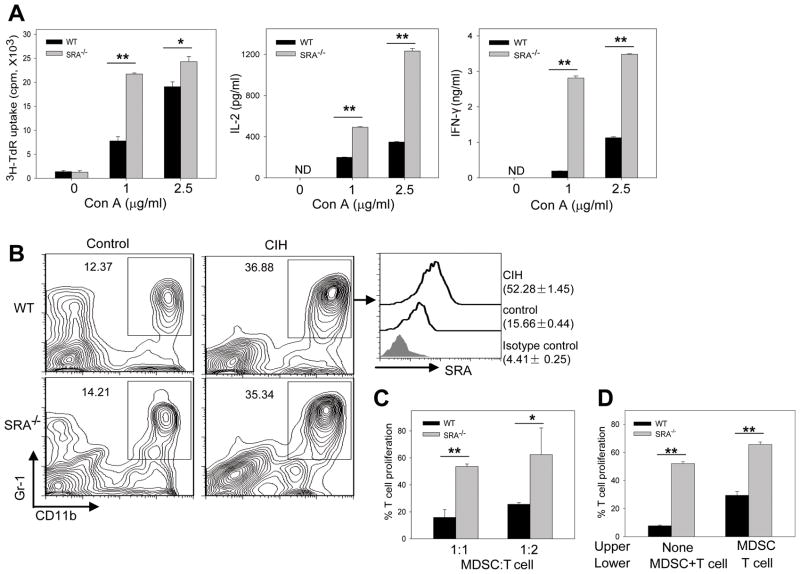
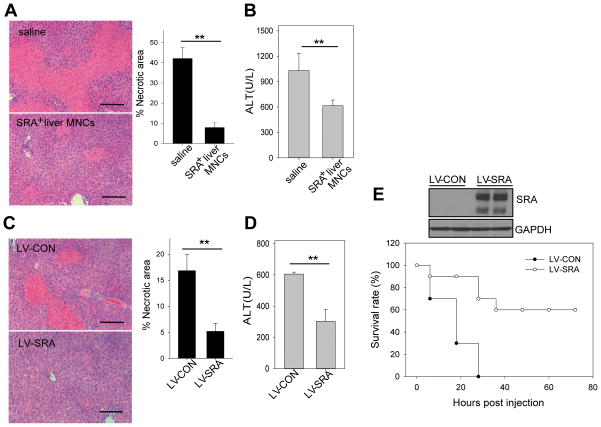
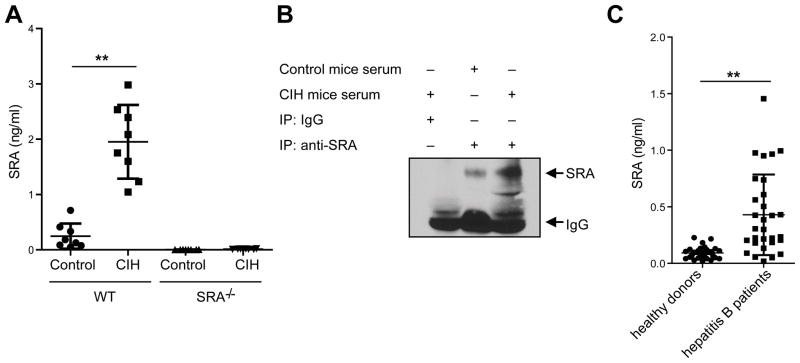
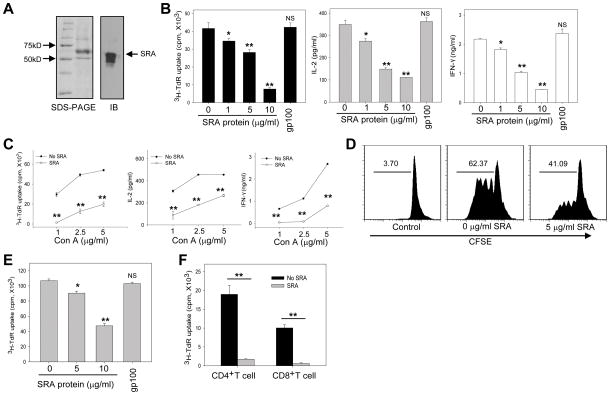
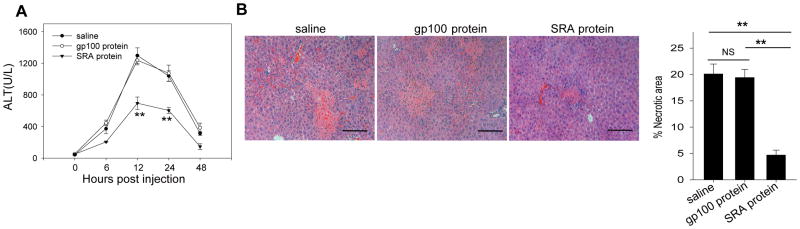
Negative feedback immune mechanisms are essential for maintenance of hepatic homeostasis and prevention of immune-mediated liver injury. We show here that scavenger receptor A (SRA/CD204), a pattern recognition molecule, is highly up-regulated in the livers of patients with autoimmune or viral hepatitis, and of mice during concanavalin A (Con A)-induced hepatitis (CIH). Strikingly, genetic SRA ablation strongly sensitizes mice to Con A-induced liver injury. SRA loss, increased mortality and liver pathology correlate with excessive production of IFN-γ and heightened activation of T cells. Increased liver expression of SRA primarily occurs in mobilized hepatic myeloid cells during CIH, including CD11b(+) Gr-1(+) cells. Mechanistic studies establish that SRA on these cells functions as a negative regulator limiting T-cell activity and cytokine production. SRA-mediated protection from CIH is further validated by adoptive transfer of SRA(+) hepatic mononuclear cells or administration of a lentivirus-expressing SRA, which effectively ameliorates Con A-induced hepatic injury. Also, CIH and clinical hepatitis are associated with increased levels of soluble SRA. This soluble SRA displays a direct T-cell inhibitory effect and is capable of mitigating Con A-induced liver pathology.
Conclusion: Our findings demonstrate an unexpected role of SRA in attenuation of Con A-induced, T-cell-mediated hepatic injury. We propose that SRA serves as an important negative feedback mechanism in liver immune homeostasis, and may be exploited for therapeutic treatment of inflammatory liver diseases.
Copyright © 2012 American Association for the Study of Liver Diseases.
Figures
References
-
- Dienes HP, Drebber U. Pathology of immune-mediated liver injury. Dig Dis. 2010;28:57–62. - PubMed
-
- Lohr HF, Schlaak JF, Gerken G, Fleischer B, Dienes HP, Meyer zum Buschenfelde KH. Phenotypical analysis and cytokine release of liver-infiltrating and peripheral blood T lymphocytes from patients with chronic hepatitis of different etiology. Liver. 1994;14:161–166. - PubMed
-
- McFarlane IG. Pathogenesis of autoimmune hepatitis. Biomed Pharmacother. 1999;53:255–263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials