

Endoplasmic reticulum stress induces a caspase-dependent N-terminal cleavage of RBX1 protein in B cells
- PMID: 22822056
- PMCID: PMC3438954
- DOI: 10.1074/jbc.M111.314583
Endoplasmic reticulum stress induces a caspase-dependent N-terminal cleavage of RBX1 protein in B cells
Abstract
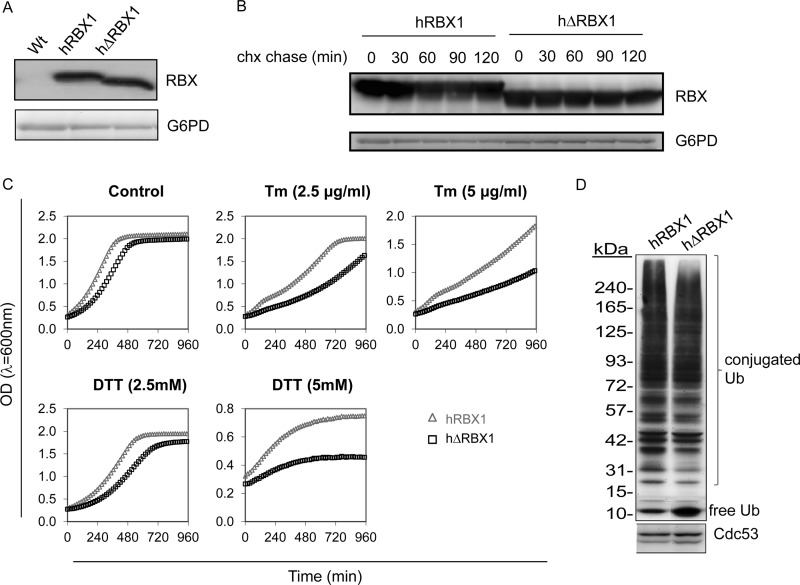
Endoplasmic reticulum (ER) stress develops when the ER is overloaded with too many proteins to fold. This elicits a signaling pathway called the unfolded protein response. The unfolded protein response is physiologically required for the terminal development of B cells into antibody-secreting plasma cells. Ring Box Protein 1 (RBX1) is a 14-kDa protein necessary for ubiquitin ligation activity of the multimeric cullin ring ubiquitin ligases (CRLs). As RBX1 is shared by a large number of CRLs, alterations in its activity may lead to global changes in protein stability. We discovered that RBX1 is cleaved in the course of LPS-induced plasma cell differentiation and in multiple myeloma cell lines upon induction of pharmacological ER stress. The cleavage is executed by several caspase proteases that cleave RBX1 eight amino acids from the N terminus. To address the possible implication of RBX1 cleavage for CRL activity, we replaced the endogenous RBX1 homolog of the yeast Saccharomyces cerevisiae, Roc1, with the wild type or the N-terminal Δ8 mutant human RBX1. We show that yeast expressing the cleaved RBX1 are hypersensitive to ER stress and are impaired in CRL-mediated ubiquitination and degradation. We propose a model by which N-terminal cleavage of RBX1 impairs its activity and promotes susceptibility to ER stress induction.
Figures




References
-
- Ron D., Walter P. (2007) Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 8, 519–529 - PubMed
-
- Travers K. J., Patil C. K., Wodicka L., Lockhart D. J., Weissman J. S., Walter P. (2000) Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 101, 249–258 - PubMed
-
- Yamamoto K., Sato T., Matsui T., Sato M., Okada T., Yoshida H., Harada A., Mori K. (2007) Transcriptional induction of mammalian ER quality control proteins is mediated by single or combined action of ATF6α and XBP1. Dev. Cell 13, 365–376 - PubMed
-
- Reimold A. M., Iwakoshi N. N., Manis J., Vallabhajosyula P., Szomolanyi-Tsuda E., Gravallese E. M., Friend D., Grusby M. J., Alt F., Glimcher L. H. (2001) Plasma cell differentiation requires the transcription factor XBP-1. Nature 412, 300–307 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
