

Sex-specific adaptation drives early sex chromosome evolution in Drosophila
- PMID: 22822149
- PMCID: PMC4107656
- DOI: 10.1126/science.1225385
Sex-specific adaptation drives early sex chromosome evolution in Drosophila
Abstract
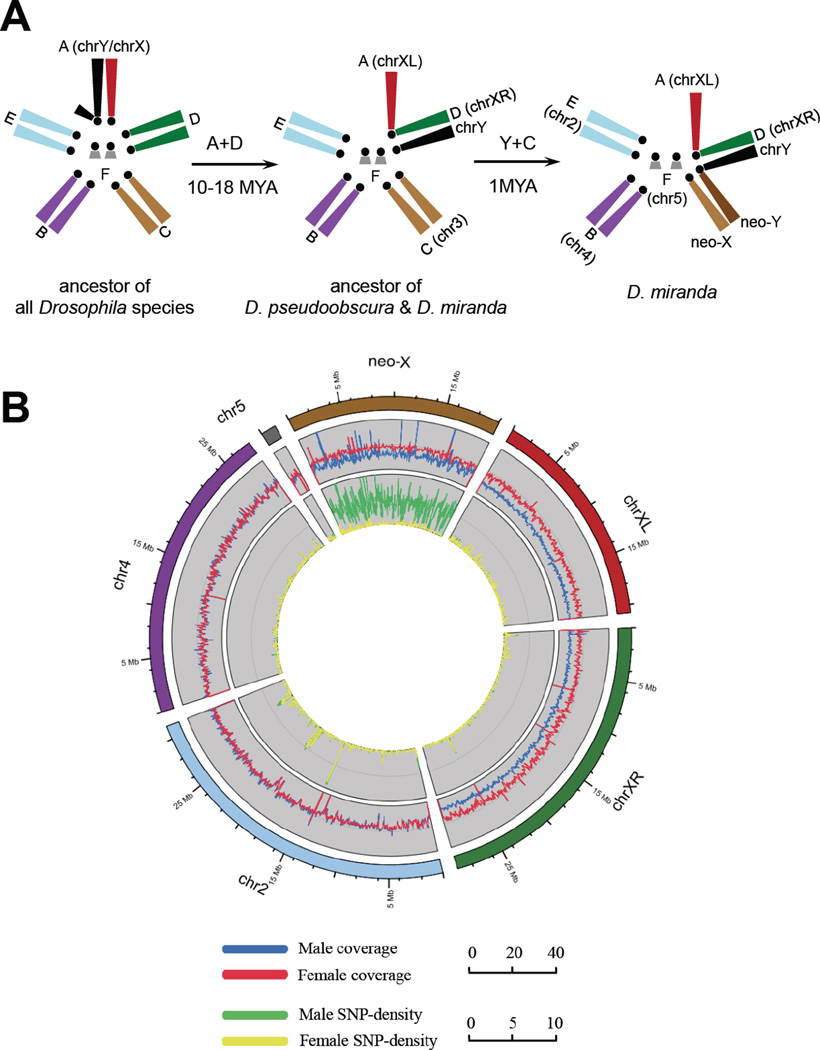
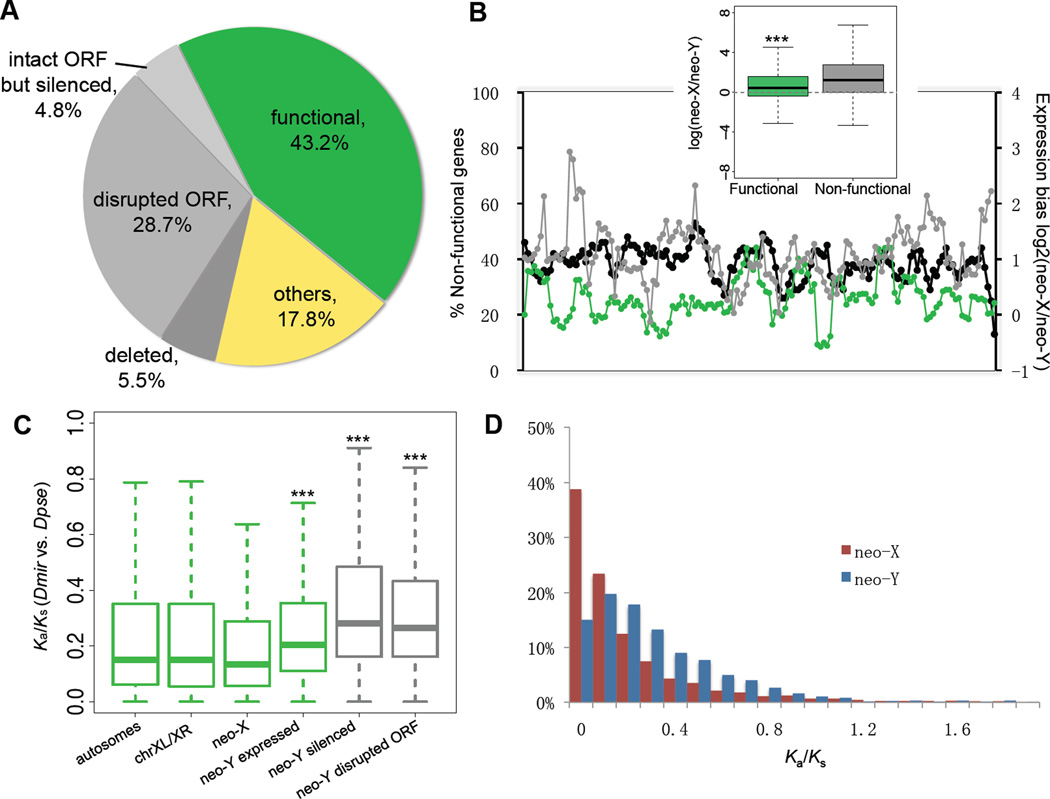
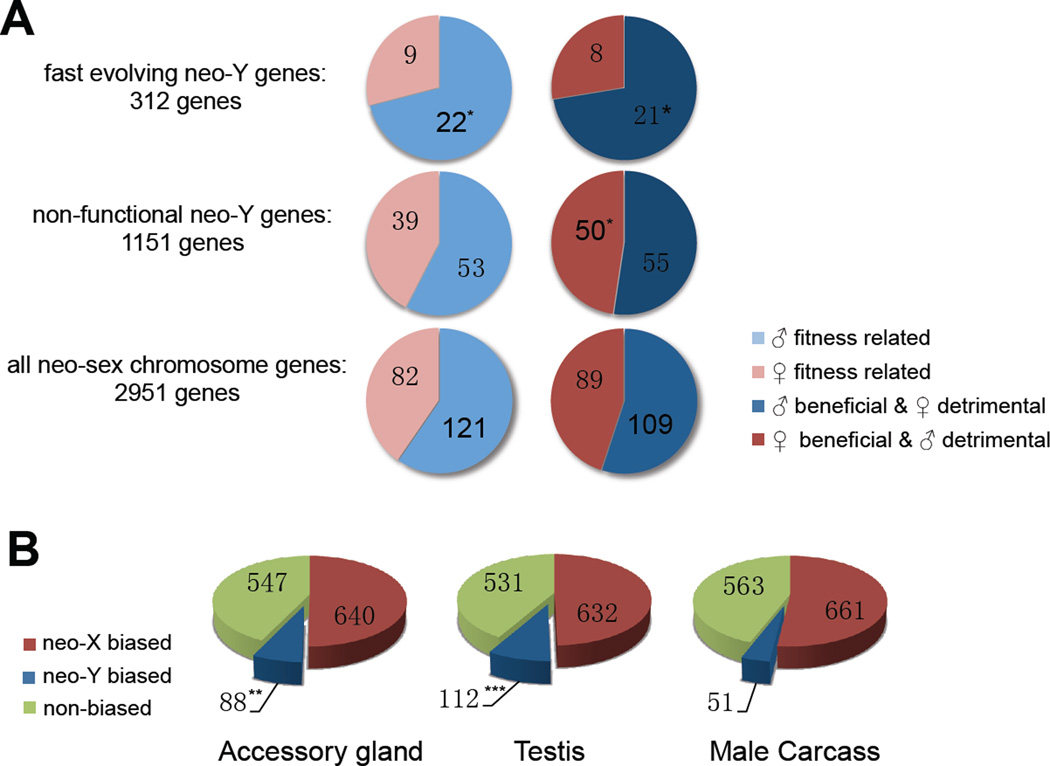
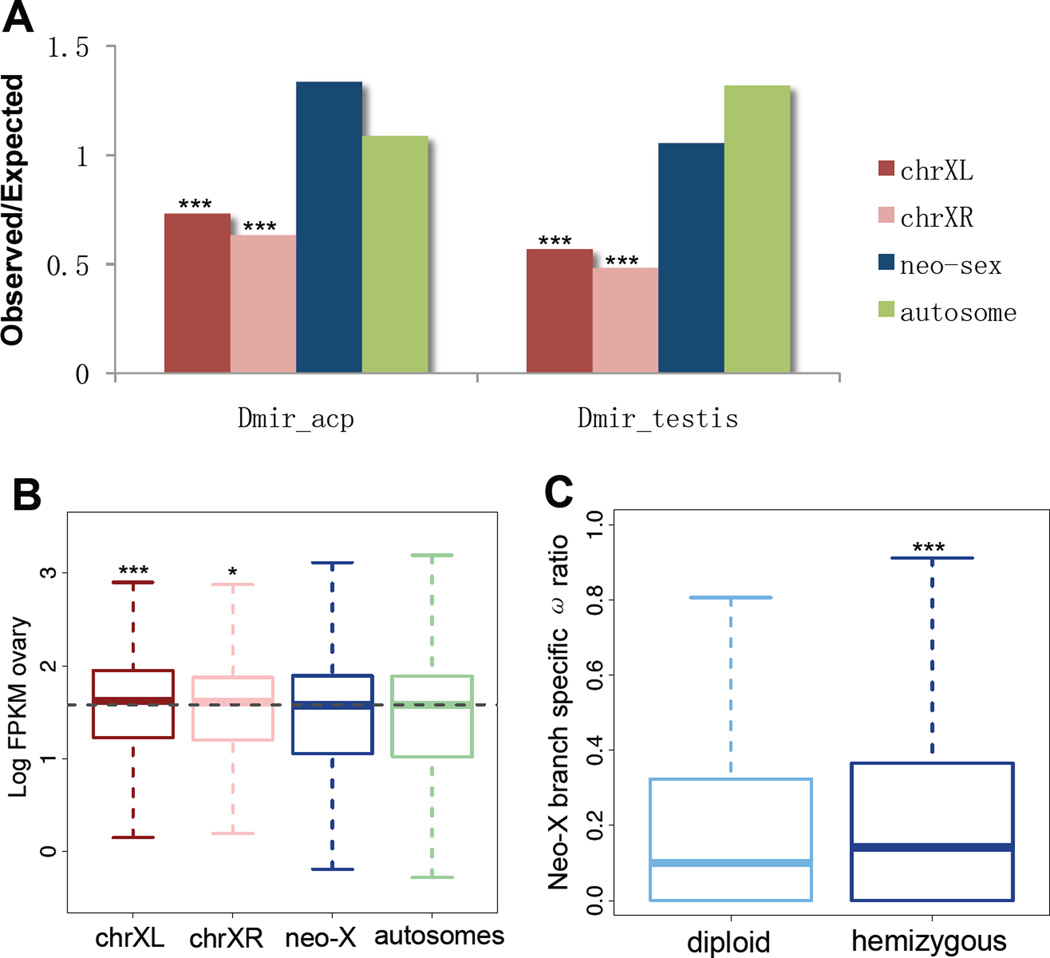
Most species' sex chromosomes are derived from ancient autosomes and show few signatures of their origins. We studied the sex chromosomes of Drosophila miranda, where a neo-Y chromosome originated only approximately 1 million years ago. Whole-genome and transcriptome analysis reveals massive degeneration of the neo-Y, that male-beneficial genes on the neo-Y are more likely to undergo accelerated protein evolution, and that neo-Y genes evolve biased expression toward male-specific tissues--the shrinking gene content of the neo-Y becomes masculinized. In contrast, although older X chromosomes show a paucity of genes expressed in male tissues, neo-X genes highly expressed in male-specific tissues undergo increased rates of protein evolution if haploid in males. Thus, the response to sex-specific selection can shift at different stages of X differentiation, resulting in masculinization or demasculinization of the X-chromosomal gene content.
Figures
References
-
- Bull JJ. Evolution of sex determining mechanisms. Benjamin Cummings; Menlo Park, CA: 1983.
-
- Skaletsky H, et al. Nature. 2003 Jun 19;423:825. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
