

Interspecific RNA interference of SHOOT MERISTEMLESS-like disrupts Cuscuta pentagona plant parasitism
- PMID: 22822208
- PMCID: PMC3426138
- DOI: 10.1105/tpc.112.099994
Interspecific RNA interference of SHOOT MERISTEMLESS-like disrupts Cuscuta pentagona plant parasitism
Abstract
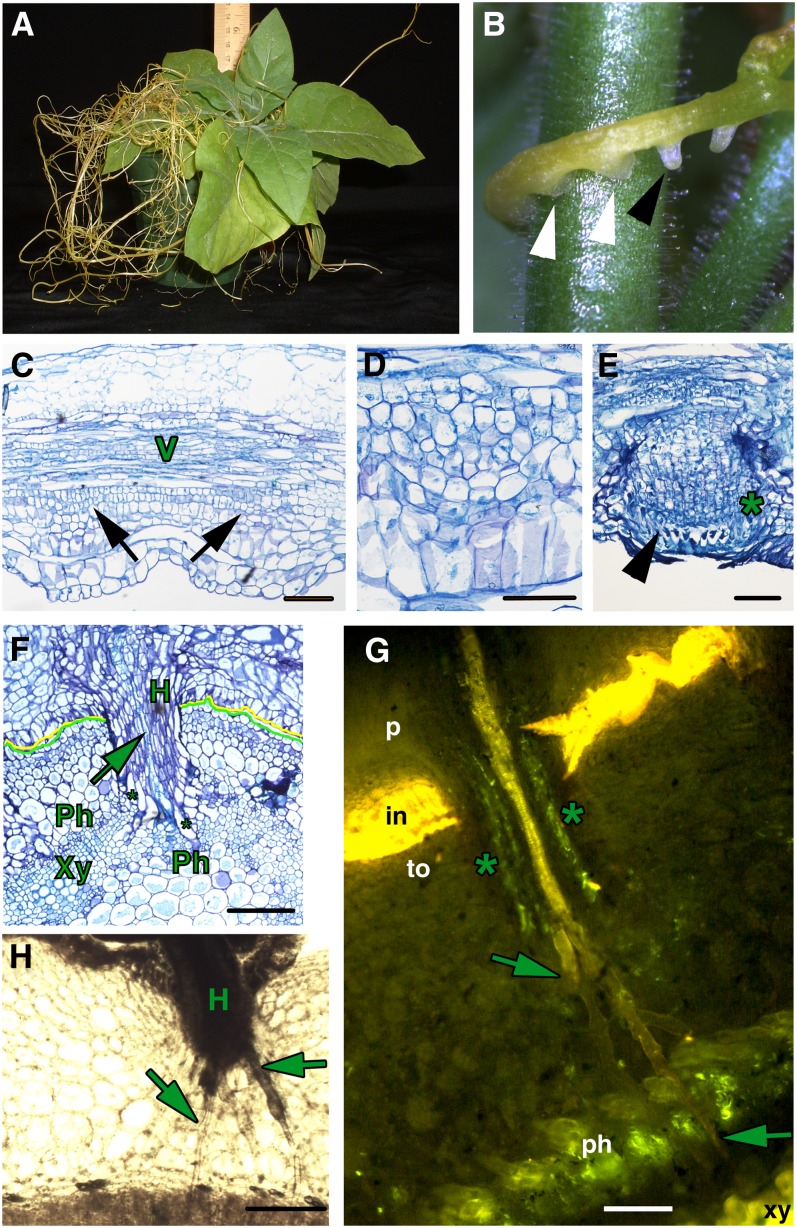
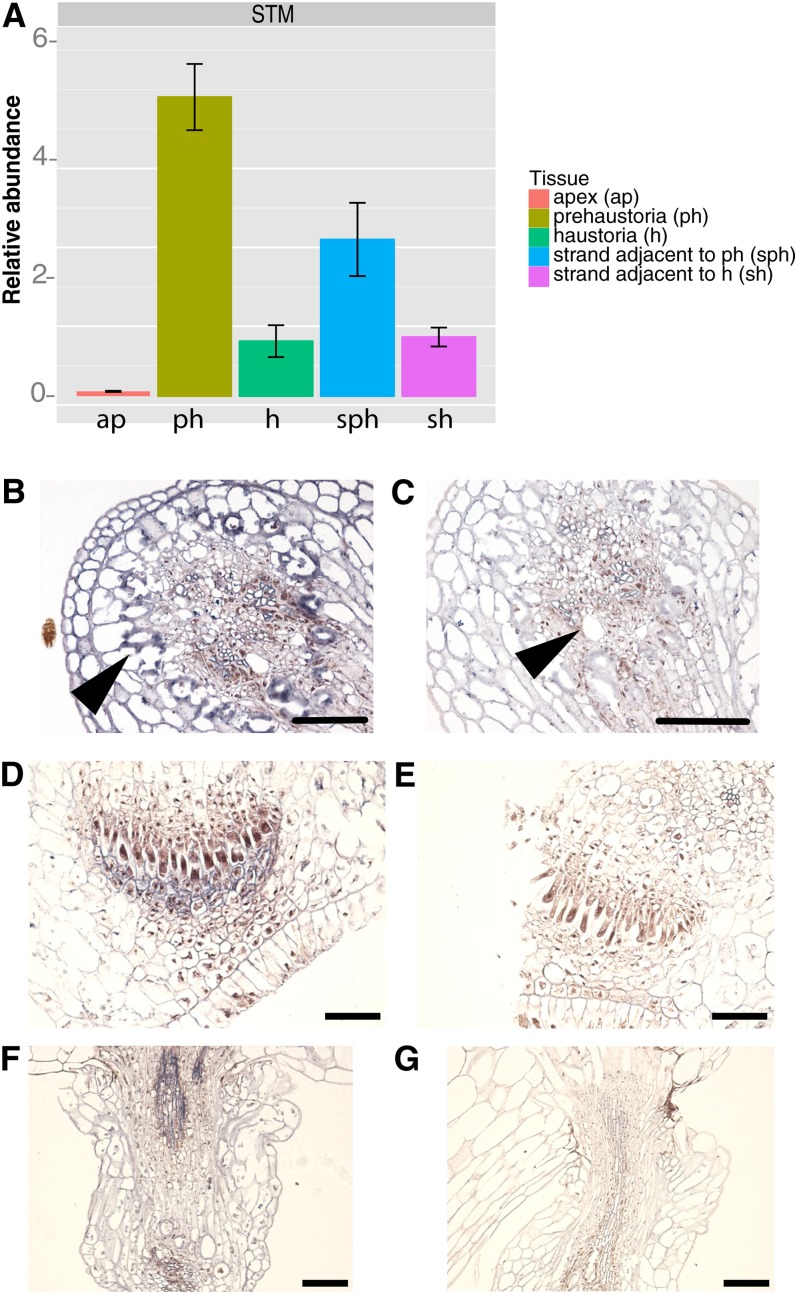
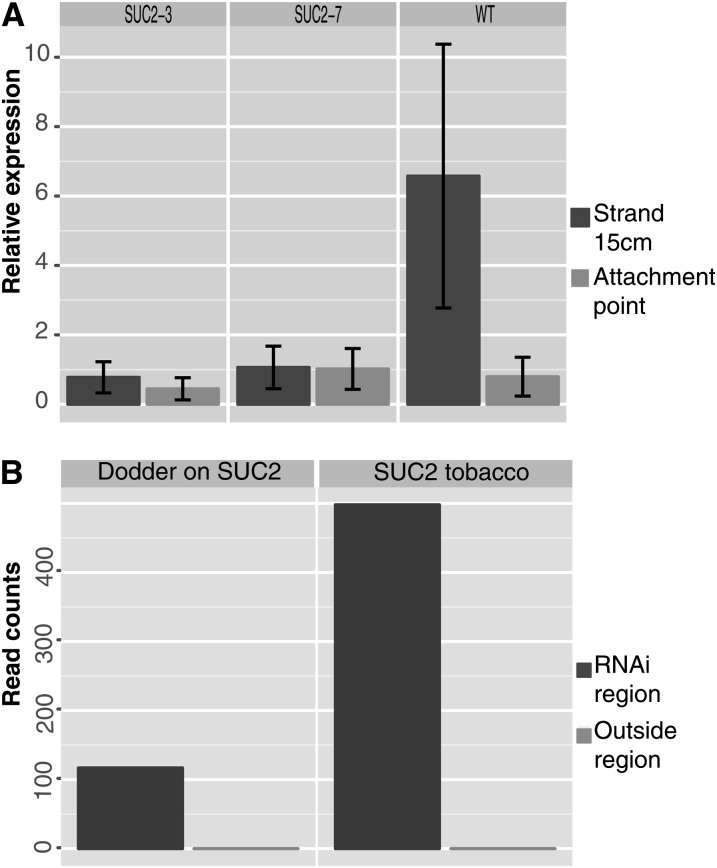
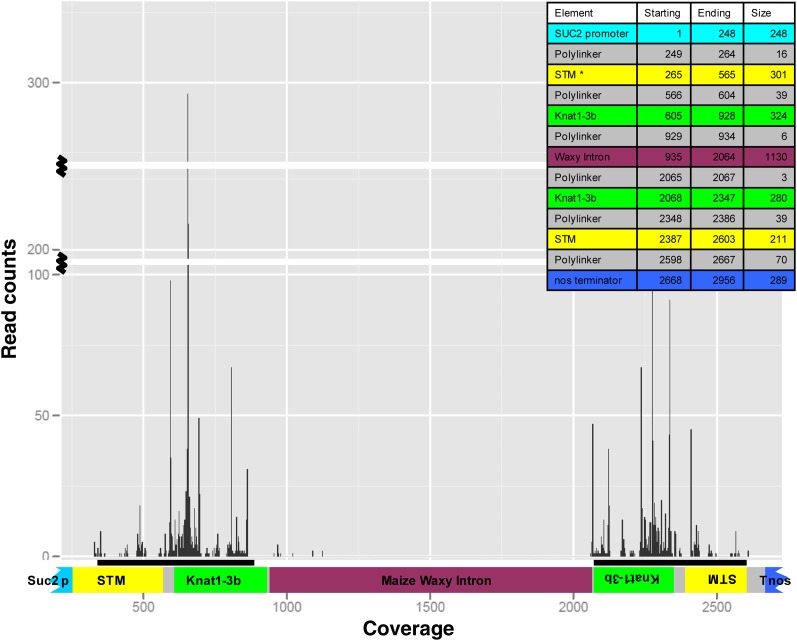
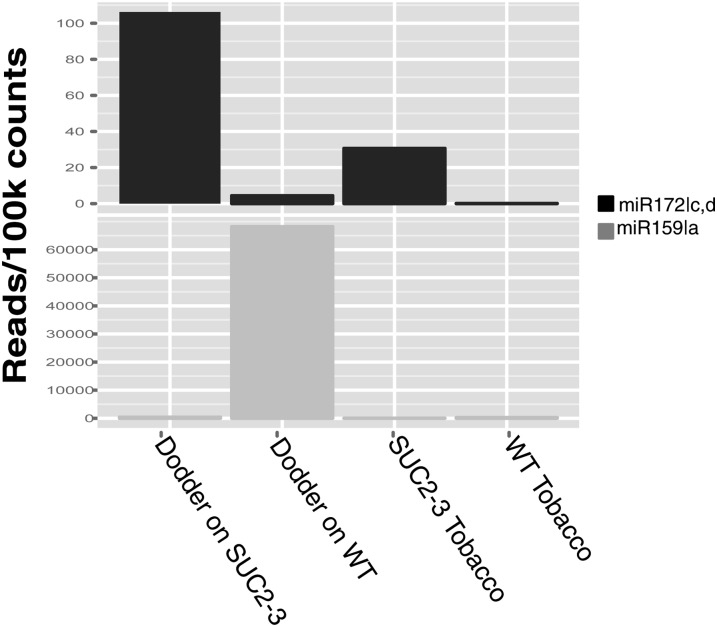
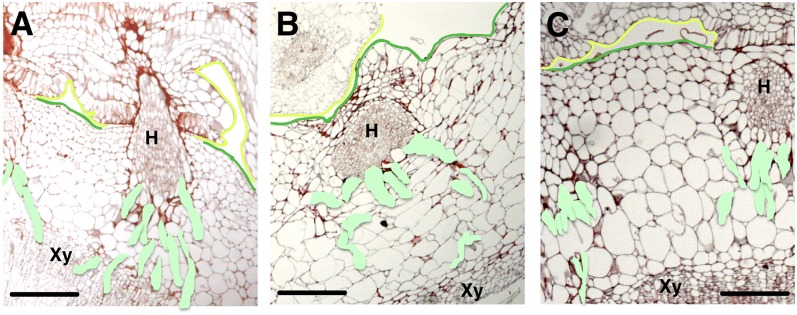
Infection of crop species by parasitic plants is a major agricultural hindrance resulting in substantial crop losses worldwide. Parasitic plants establish vascular connections with the host plant via structures termed haustoria, which allow acquisition of water and nutrients, often to the detriment of the infected host. Despite the agricultural impact of parasitic plants, the molecular and developmental processes by which host/parasitic interactions are established are not well understood. Here, we examine the development and subsequent establishment of haustorial connections by the parasite dodder (Cuscuta pentagona) on tobacco (Nicotiana tabacum) plants. Formation of haustoria in dodder is accompanied by upregulation of dodder KNOTTED-like homeobox transcription factors, including SHOOT MERISTEMLESS-like (STM). We demonstrate interspecific silencing of a STM gene in dodder driven by a vascular-specific promoter in transgenic host plants and find that this silencing disrupts dodder growth. The reduced efficacy of dodder infection on STM RNA interference transgenics results from defects in haustorial connection, development, and establishment. Identification of transgene-specific small RNAs in the parasite, coupled with reduced parasite fecundity and increased growth of the infected host, demonstrates the efficacy of interspecific small RNA-mediated silencing of parasite genes. This technology has the potential to be an effective method of biological control of plant parasite infection.
Figures

Comment in
-
No more free lunch: using RNA interference in the host to reduce growth of a parasitic plant.Plant Cell. 2012 Jul;24(7):2709. doi: 10.1105/tpc.112.240712. Epub 2012 Jul 24. Plant Cell. 2012. PMID: 22829144 Free PMC article. No abstract available.
References
-
- Abraham-Juárez M.J., Martínez-Hernández A., Leyva-González M.A., Herrera-Estrella L., Simpson J. (2010). Class I KNOX genes are associated with organogenesis during bulbil formation in Agave tequilana. J. Exp. Bot. 61: 4055–4067 - PubMed
-
- Achard P., Herr A., Baulcombe D.C., Harberd N.P. (2004). Modulation of floral development by a gibberellin-regulated microRNA. Development 131: 3357–3365 - PubMed
-
- Ahkami A.H., Lischewski S., Haensch K.T., Porfirova S., Hofmann J., Rolletschek H., Melzer M., Franken P., Hause B., Druege U., Hajirezaei M.R. (2009). Molecular physiology of adventitious root formation in Petunia hybrida cuttings: Involvement of wound response and primary metabolism. New Phytol. 181: 613–625 - PubMed
-
- Aly R., Cholakh H., Joel D.M., Leibman D., Steinitz B., Zelcer A., Naglis A., Yarden O., Gal-On A. (2009). Gene silencing of mannose 6-phosphate reductase in the parasitic weed Orobanche aegyptiaca through the production of homologous dsRNA sequences in the host plant. Plant Biotechnol. J. 7: 487–498 - PubMed
-
- Aly R., Hamamouch N., Abu-Nassar J., Wolf S., Joel D.M., Eizenberg H., Kaisler E., Cramer C., Gal-On A., Westwood J.H. (2011). Movement of protein and macromolecules between host plants and the parasitic weed Phelipanche aegyptiaca Pers. Plant Cell Rep. 30: 2233–2241 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
