

Phosphorylation primes vinculin for activation
- PMID: 22824265
- PMCID: PMC3341567
- DOI: 10.1016/j.bpj.2012.01.062
Phosphorylation primes vinculin for activation
Abstract
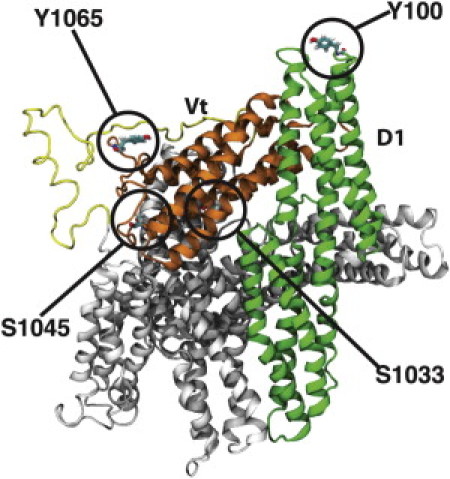
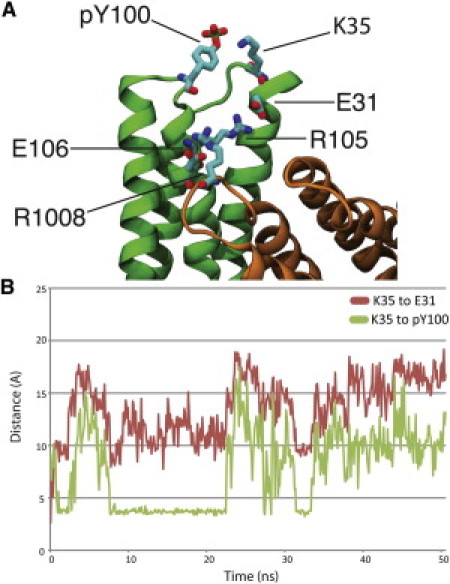
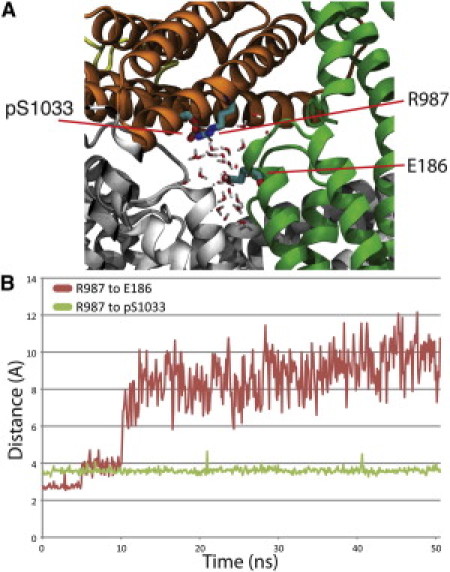
Vinculin phosphorylation has been implicated as a potential mechanism for focal adhesion growth and maturation. Four vinculin residues-Y100, S1033, S1045, and Y1065-are phosphorylated by kinases during focal adhesion maturation. In this study, phosphorylation at each of these residues is simulated using molecular dynamics models. The simulations demonstrate that once each phosphorylated vinculin structure is at equilibrium, significant local conformational changes result that may impact either vinculin activation or vinculin binding to actin and PIP2. Simulation of vinculin activation after phosphorylation shows that the added phosphoryl groups can prime vinculin for activation. It remains to be seen if vinculin can be phosphorylated at S1033 in vivo, but these simulations highlight that in the event of a S1033 phophorylation vinculin will likely be primed for activation.
Copyright © 2012 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures




Similar articles
-
Molecular dynamics study of talin-vinculin binding.Biophys J. 2008 Aug;95(4):2027-36. doi: 10.1529/biophysj.107.124487. Epub 2008 Apr 11. Biophys J. 2008. PMID: 18408041 Free PMC article.
-
How force might activate talin's vinculin binding sites: SMD reveals a structural mechanism.PLoS Comput Biol. 2008 Feb;4(2):e24. doi: 10.1371/journal.pcbi.0040024. PLoS Comput Biol. 2008. PMID: 18282082 Free PMC article.
-
Vinculin phosphorylation at residues Y100 and Y1065 is required for cellular force transmission.J Cell Sci. 2015 Sep 15;128(18):3435-43. doi: 10.1242/jcs.172031. Epub 2015 Aug 3. J Cell Sci. 2015. PMID: 26240176 Free PMC article.
-
Cytoskeletal proteins talin and vinculin in integrin-mediated adhesion.Biochem Soc Trans. 2004 Nov;32(Pt 5):831-6. doi: 10.1042/BST0320831. Biochem Soc Trans. 2004. PMID: 15494027 Review.
-
The structure and regulation of vinculin.Trends Cell Biol. 2006 Sep;16(9):453-60. doi: 10.1016/j.tcb.2006.07.004. Epub 2006 Aug 8. Trends Cell Biol. 2006. PMID: 16893648 Review.
Cited by
-
The interaction of vinculin with actin.PLoS Comput Biol. 2013 Apr;9(4):e1002995. doi: 10.1371/journal.pcbi.1002995. Epub 2013 Apr 25. PLoS Comput Biol. 2013. PMID: 23633939 Free PMC article.
-
Vinculin phosphorylation at Tyr1065 regulates vinculin conformation and tension development in airway smooth muscle tissues.J Biol Chem. 2014 Feb 7;289(6):3677-88. doi: 10.1074/jbc.M113.508077. Epub 2013 Dec 13. J Biol Chem. 2014. PMID: 24338477 Free PMC article.
-
Shp2 plays a crucial role in cell structural orientation and force polarity in response to matrix rigidity.Proc Natl Acad Sci U S A. 2013 Feb 19;110(8):2840-5. doi: 10.1073/pnas.1222164110. Epub 2013 Jan 28. Proc Natl Acad Sci U S A. 2013. PMID: 23359696 Free PMC article.
-
Pulmonary Endothelial Mechanical Sensing and Signaling, a Story of Focal Adhesions and Integrins in Ventilator Induced Lung Injury.Front Physiol. 2019 Apr 26;10:511. doi: 10.3389/fphys.2019.00511. eCollection 2019. Front Physiol. 2019. PMID: 31105595 Free PMC article. Review.
-
The talin dimer structure orientation is mechanically regulated.Biophys J. 2014 Oct 21;107(8):1802-1809. doi: 10.1016/j.bpj.2014.08.038. Biophys J. 2014. PMID: 25418161 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
