

Degradation of endocytosed gap junctions by autophagosomal and endo-/lysosomal pathways: a perspective
- PMID: 22825714
- PMCID: PMC3458158
- DOI: 10.1007/s00232-012-9464-0
Degradation of endocytosed gap junctions by autophagosomal and endo-/lysosomal pathways: a perspective
Abstract
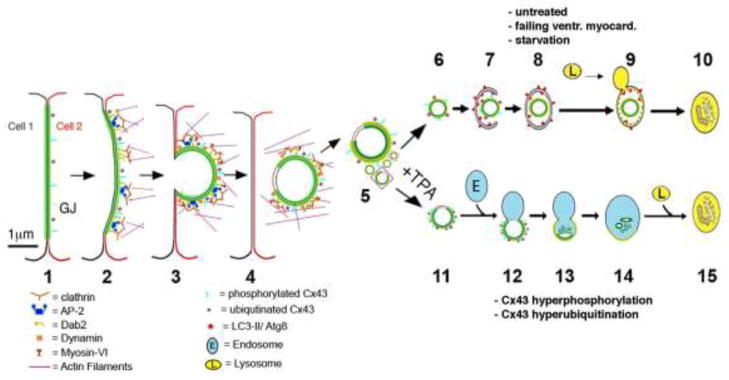
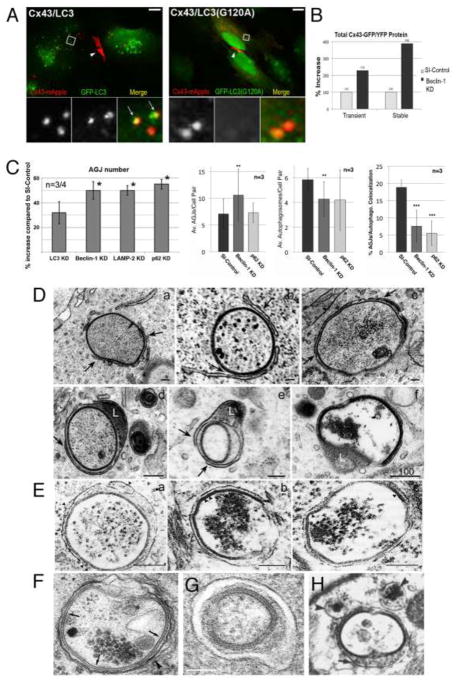
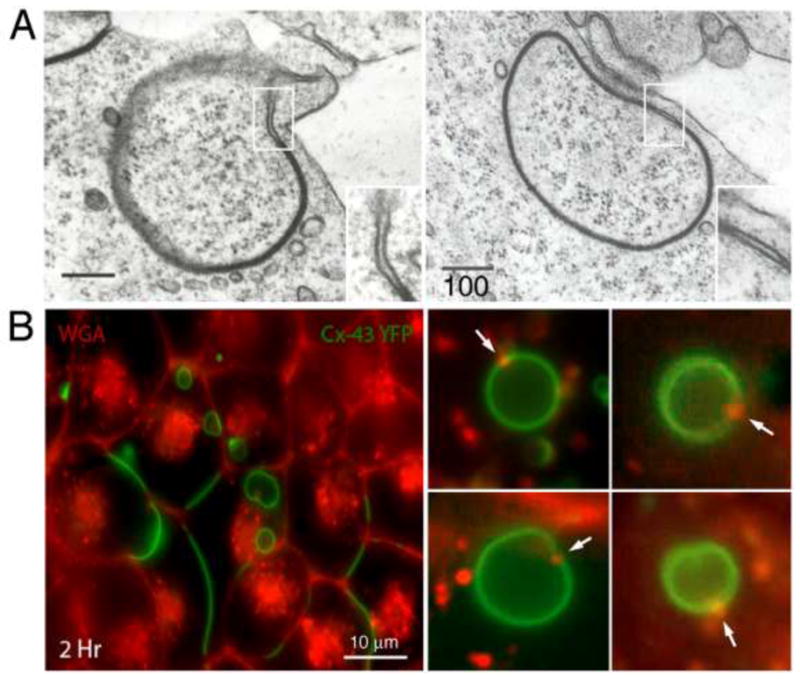
Gap junctions (GJs) are composed of tens to many thousands of double-membrane spanning GJ channels that cluster together to form densely packed channel arrays (termed GJ plaques) in apposing plasma membranes of neighboring cells. In addition to providing direct intercellular communication (GJIC, their hallmark function), GJs, based on their characteristic double-membrane-spanning configuration, likely also significantly contribute to physical cell-to-cell adhesion. Clearly, modulation (up-/down-regulation) of GJIC and of physical cell-to-cell adhesion is as vitally important as the basic ability of GJ formation itself. Others and we have previously described that GJs can be removed from the plasma membrane via the internalization of entire GJ plaques (or portions thereof) in a cellular process that resembles clathrin-mediated endocytosis. GJ endocytosis results in the formation of double-membrane vesicles [termed annular gap junctions (AGJs) or connexosomes] in the cytoplasm of one of the coupled cells. Four recent independent studies, consistent with earlier ultrastructural analyses, demonstrate the degradation of endocytosed AGJ vesicles via autophagy. However, in TPA-treated cells others report degradation of AGJs via the endo-/lysosomal degradation pathway. Here we summarize evidence that supports the concept that autophagy serves as the cellular default pathway for the degradation of internalized GJs. Furthermore, we highlight and discuss structural criteria that seem required for an alternate degradation via the endo-/lysosomal pathway.
Figures
References
-
- Baldwin AL, Thurston G. Mechanics of endothelial cell architecture and vascular permeability. Crit Rev Biomed Eng. 2001;29:247–278. - PubMed
-
- Barriere H, Nemes C, Lechardeur D, Khan-Mohammad M, Fruh K, Lukacs GL. Molecular basis of oligoubiquitin-dependent internalization of membrane proteins in Mammalian cells. Traffic. 2006;7:282–297. - PubMed
-
- Beardslee MA, Laing JG, Beyer EC, Saffitz JE. Rapid turnover of connexin43 in the adult rat heart. Circ Res. 1998;83:629–635. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
