

Inhibition of mechanistic target of rapamycin promotes dendritic cell activation and enhances therapeutic autologous vaccination in mice
- PMID: 22826320
- PMCID: PMC3424310
- DOI: 10.4049/jimmunol.1103741
Inhibition of mechanistic target of rapamycin promotes dendritic cell activation and enhances therapeutic autologous vaccination in mice
Abstract
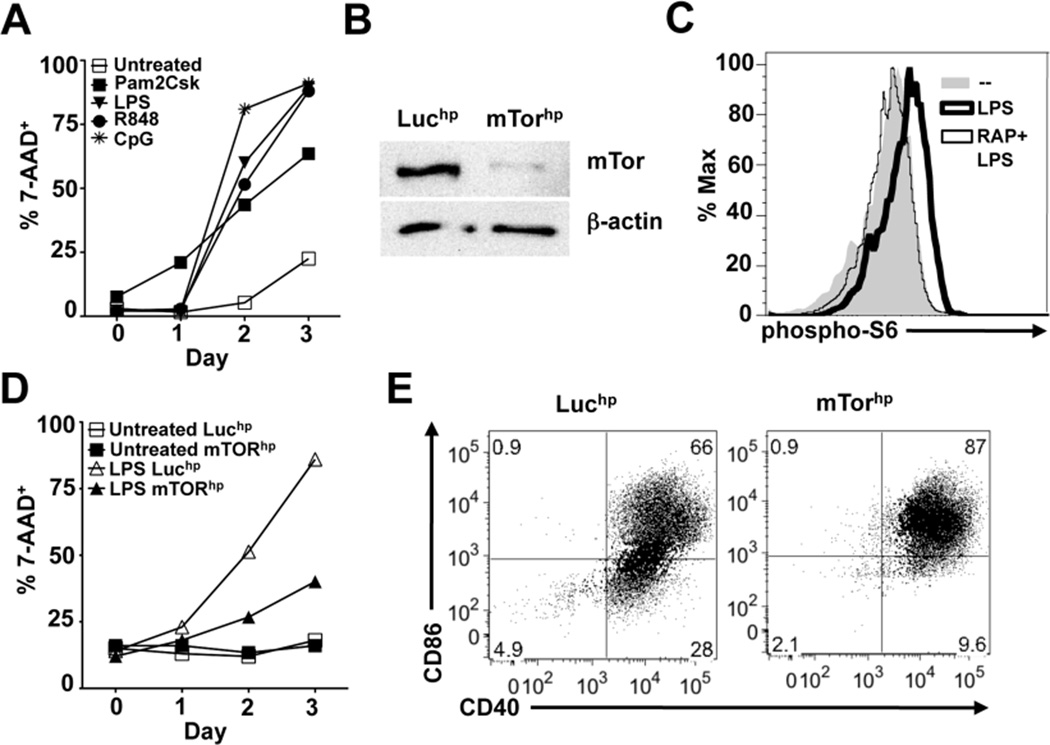
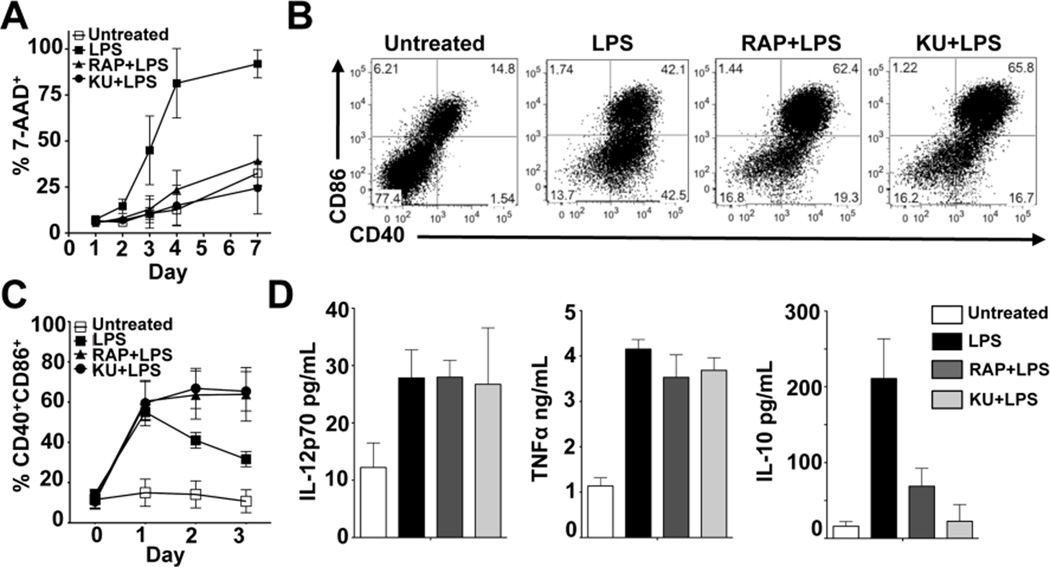
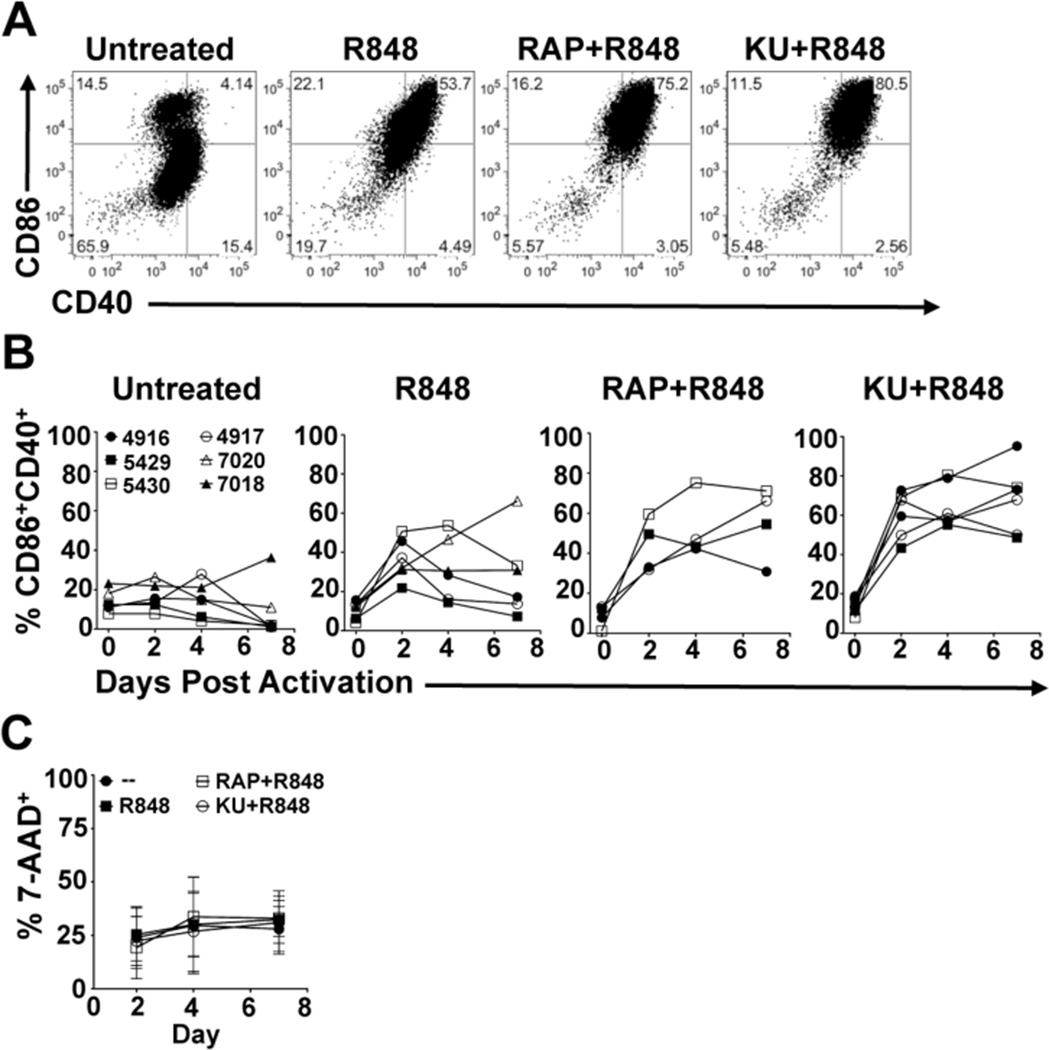
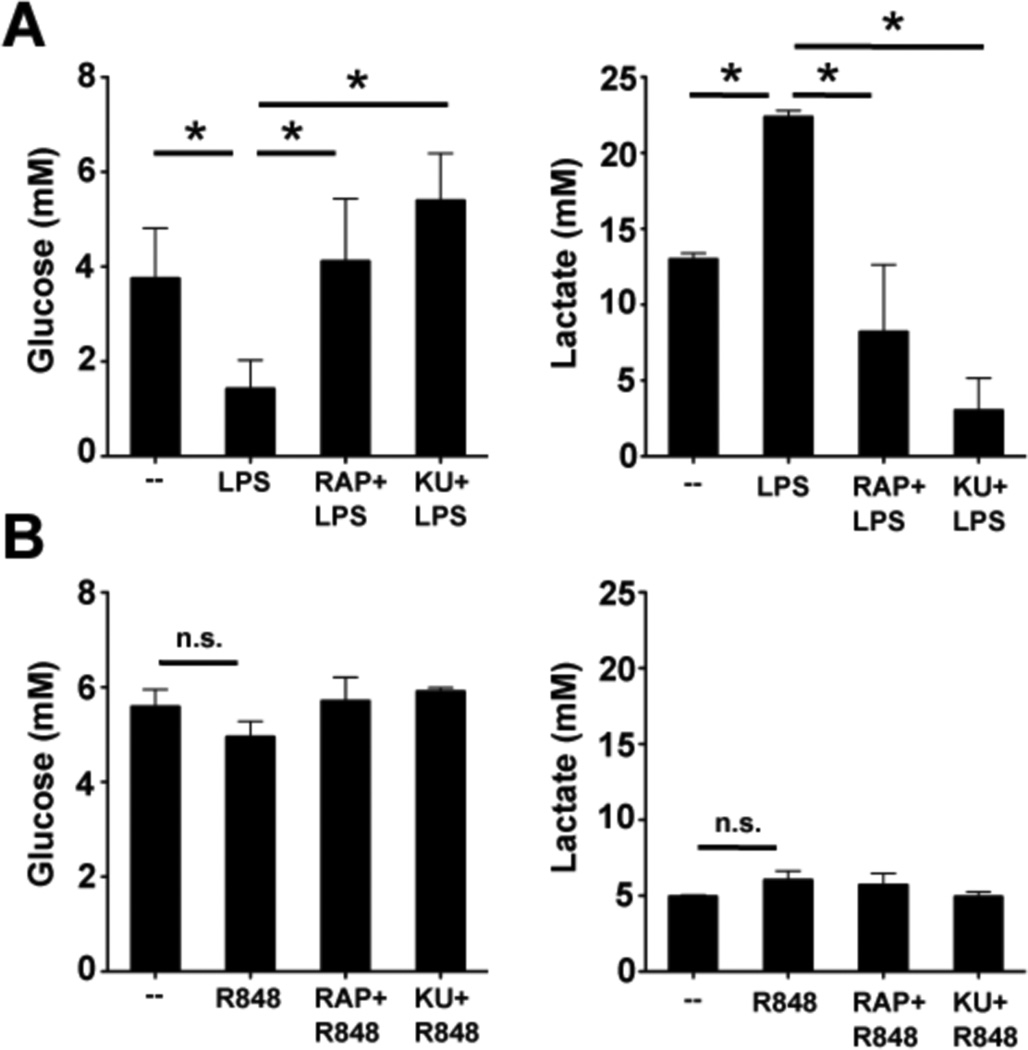
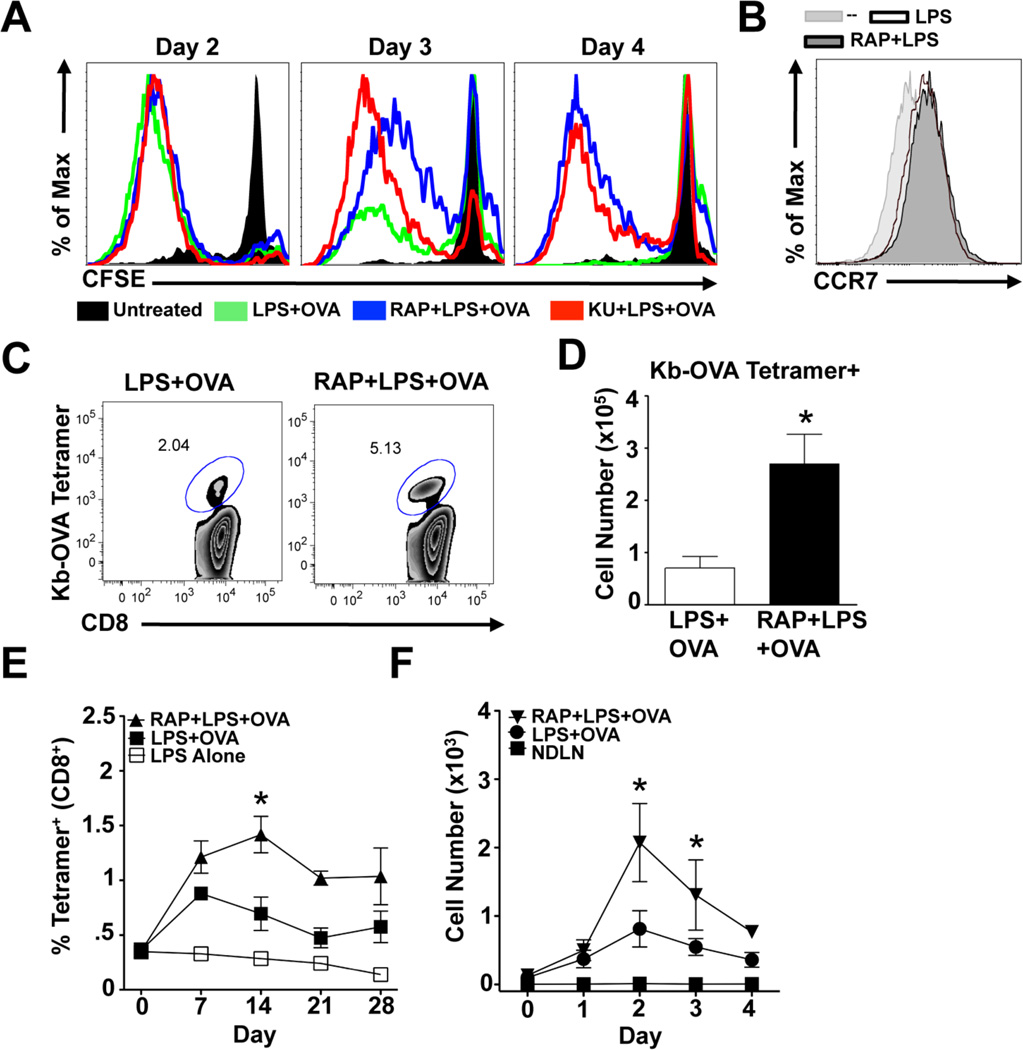
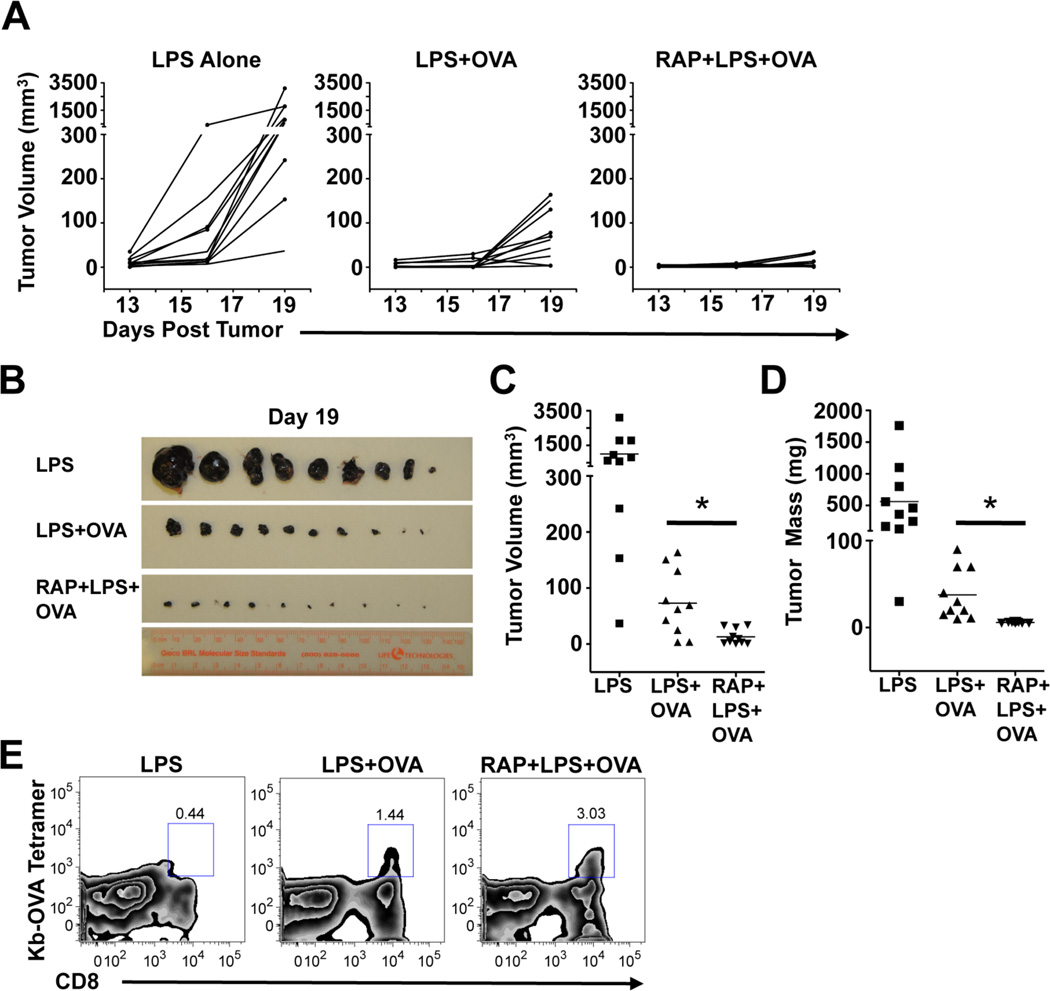
Dendritic cells (DCs) are potent inducers of T cell immunity, and autologous DC vaccination holds promise for the treatment of cancers and chronic infectious diseases. In practice, however, therapeutic vaccines of this type have had mixed success. In this article, we show that brief exposure to inhibitors of mechanistic target of rapamycin (mTOR) in DCs during the period that they are responding to TLR agonists makes them particularly potent activators of naive CD8+ T cells and able to enhance control of B16 melanoma in a therapeutic autologous vaccination model in the mouse. The improved performance of DCs in which mTOR has been inhibited is correlated with an extended life span after activation and prolonged, increased expression of costimulatory molecules. Therapeutic autologous vaccination with DCs treated with TLR agonists plus the mTOR inhibitor rapamycin results in improved generation of Ag-specific CD8+ T cells in vivo and improved antitumor immunity compared with that observed with DCs treated with TLR agonists alone. These findings define mTOR as a molecular target for augmenting DC survival and activation, and document a novel pharmacologic approach for enhancing the efficacy of therapeutic autologous DC vaccination.
Figures
References
-
- Barton GM, Medzhitov R. Control of adaptive immune responses by Toll-like receptors. Curr Opin Immunol. 2002;14:380–383. - PubMed
-
- Steinman RM, Hemmi H. Dendritic cells: translating innate to adaptive immunity. Curr Top Microbiol Immunol. 2006;311:17–58. - PubMed
-
- Palucka AK, Ueno H, Fay JW, Banchereau J. Taming cancer by inducing immunity via dendritic cells. Immunol Rev. 2007;220:129–150. - PubMed
-
- Kim JH, Kang TH, Noh KH, Bae HC, Kim SH, Yoo YD, Seong SY, Kim TW. Enhancement of dendritic cell-based vaccine potency by anti-apoptotic siRNAs targeting key pro-apoptotic proteins in cytotoxic CD8(+) T cell-mediated cell death. Immunol Lett. 2009;122:58–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
