

Systematic, balancing gradients in neuron density and number across the primate isocortex
- PMID: 22826696
- PMCID: PMC3399120
- DOI: 10.3389/fnana.2012.00028
Systematic, balancing gradients in neuron density and number across the primate isocortex
Abstract
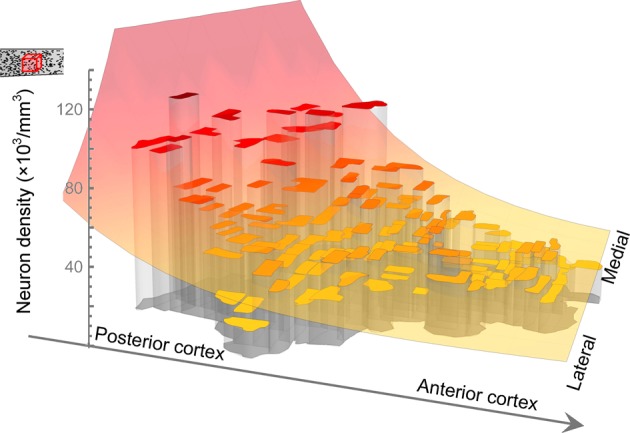
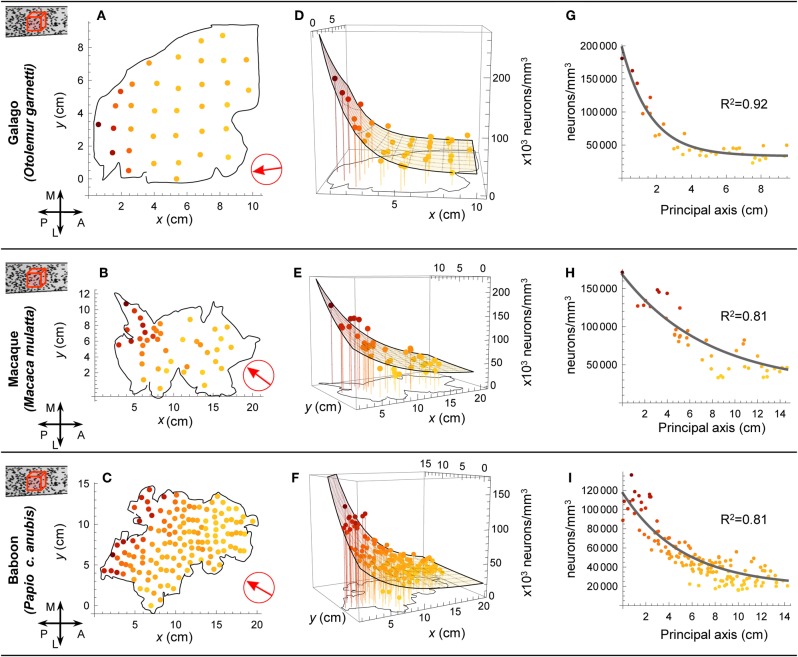
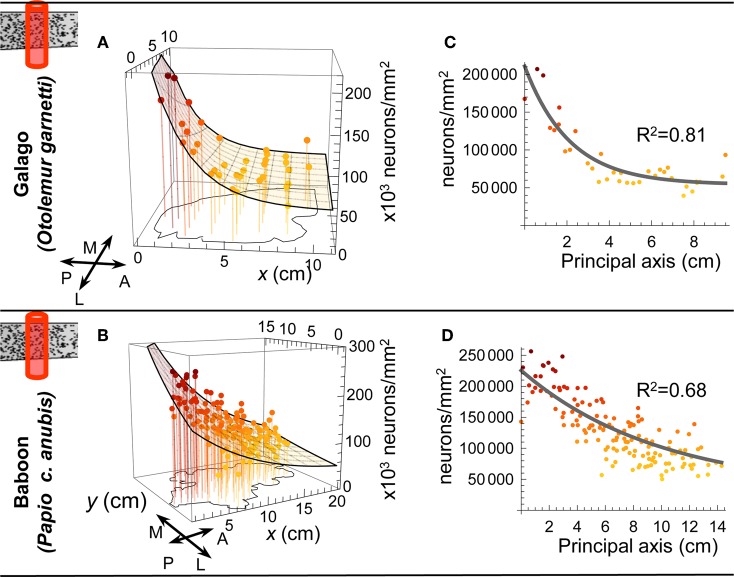
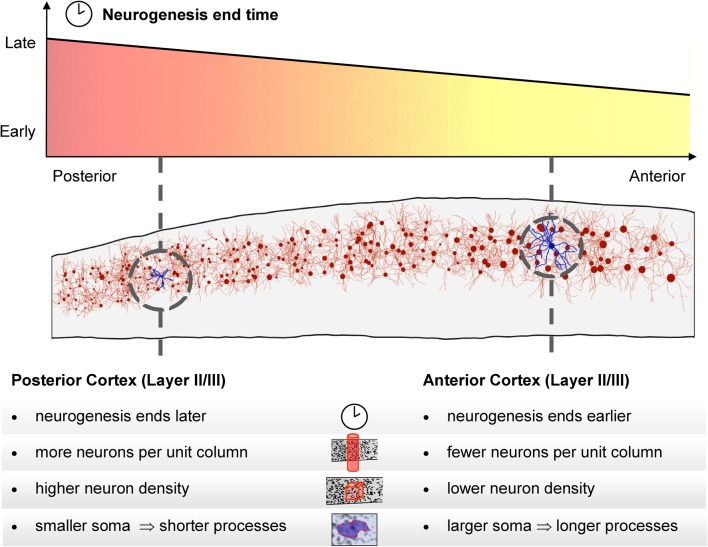
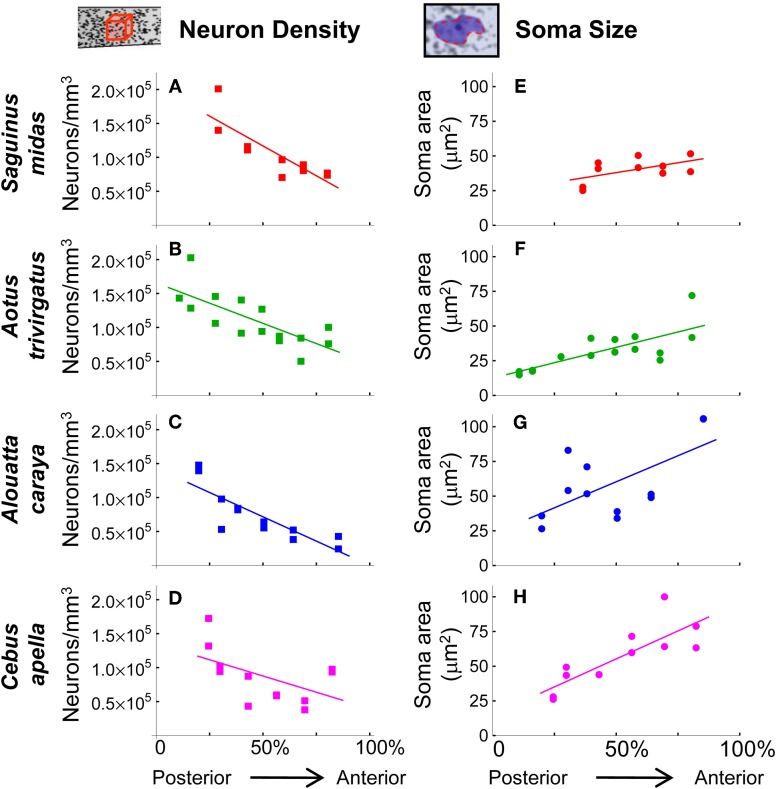
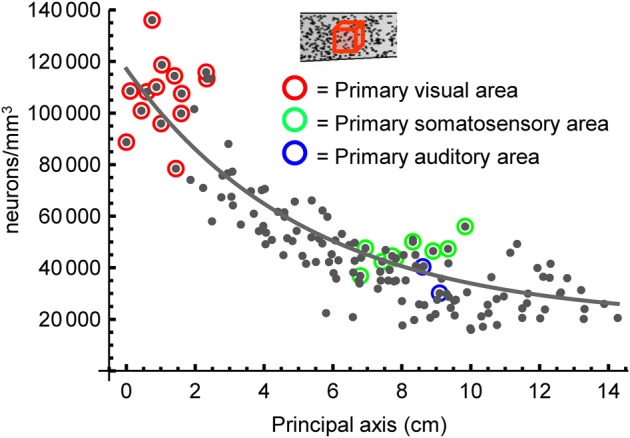
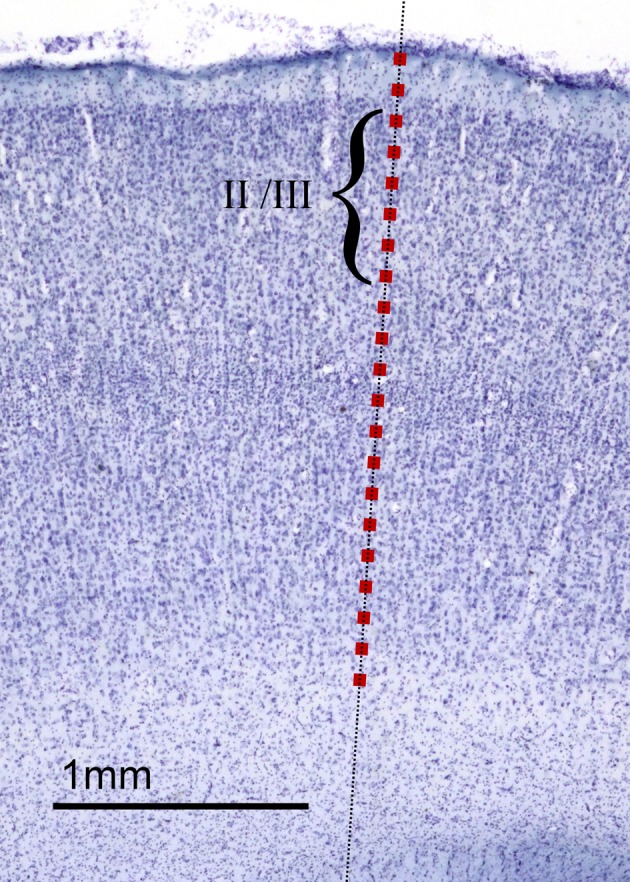
The cellular and areal organization of the cerebral cortex impacts how it processes and integrates information. How that organization emerges and how best to characterize it has been debated for over a century. Here we demonstrate and describe in the isocortices of seven primate species a pronounced anterior-to-posterior gradient in the density of neurons and in the number of neurons under a unit area of the cortical surface. Our findings assert that the cellular architecture of the primate isocortex is neither arranged uniformly nor into discrete patches with an arbitrary spatial arrangement. Rather, it exhibits striking systematic variation. We conjecture that these gradients, which establish the basic landscape that richer areal and cellular structure is built upon, result from developmental patterns of cortical neurogenesis which are conserved across species. Moreover, we propose a functional consequence: that the gradient in neurons per unit of cortical area fosters the integration and dimensional reduction of information along its ascent through sensory areas and toward frontal cortex.
Keywords: cortex; cortical areas; cytoarchitecture; evolutionary development; gradient; neurogenesis; primate evolution.
Figures

References
-
- Bayer S. A., Altman J. (1991). Neocortical development. New York, NY: Raven Press;
Grants and funding
LinkOut - more resources
Full Text Sources