

Oxygen consumption and distribution in the Long-Evans rat retina
- PMID: 22828049
- PMCID: PMC3437263
- DOI: 10.1016/j.exer.2012.07.004
Oxygen consumption and distribution in the Long-Evans rat retina
Abstract
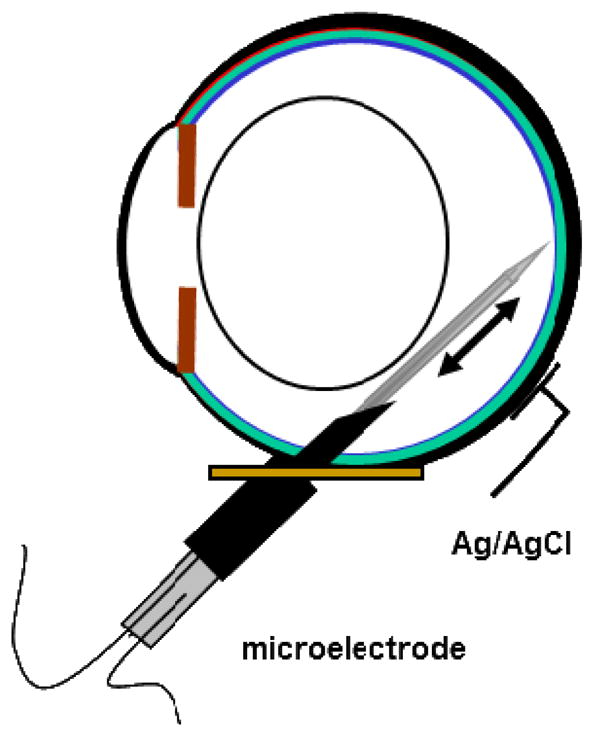
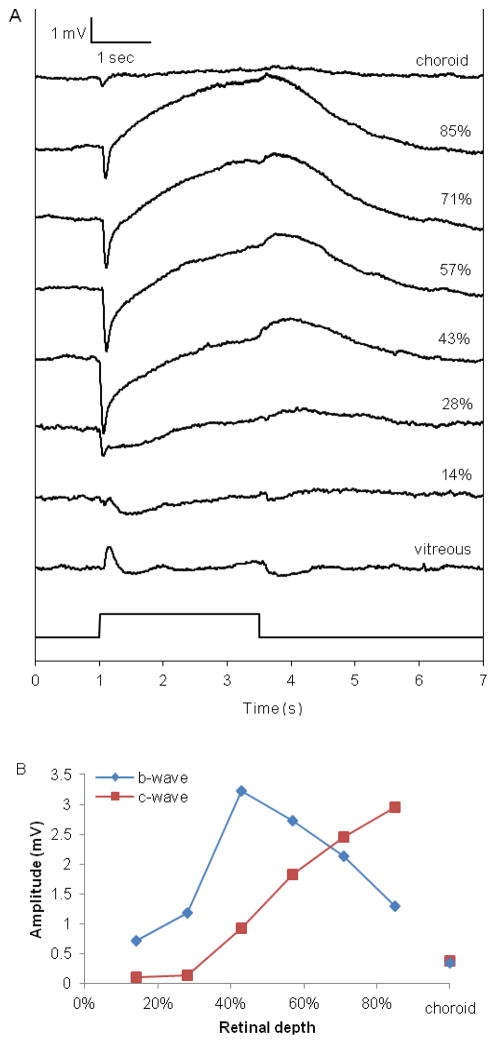
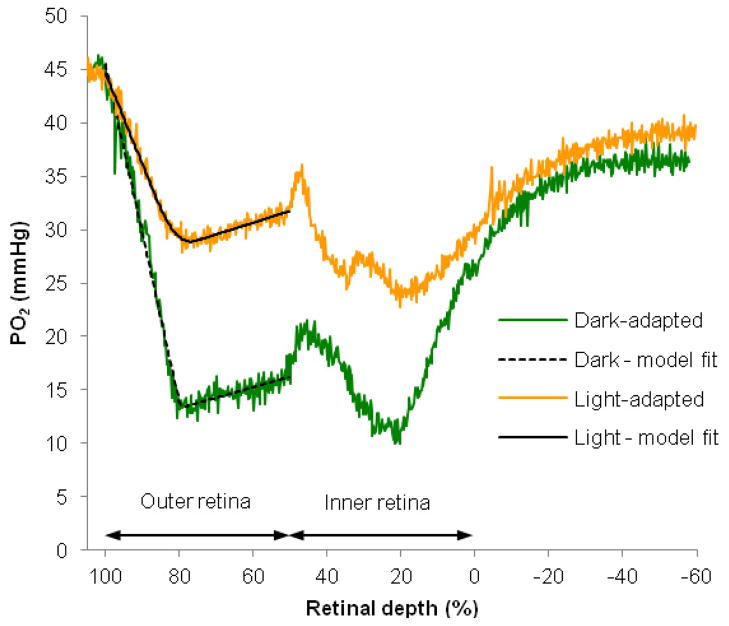
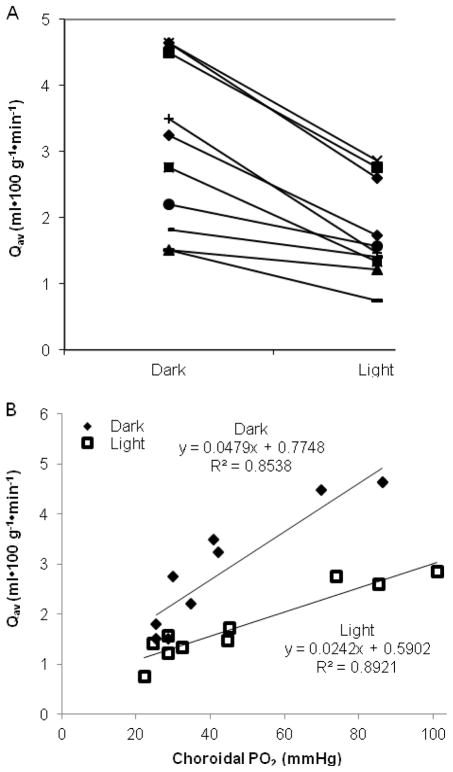
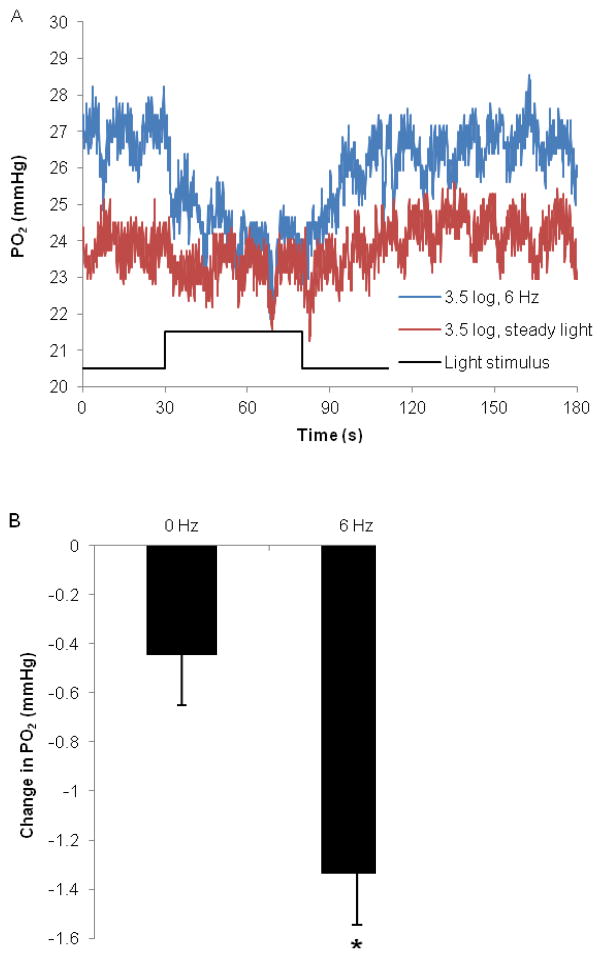
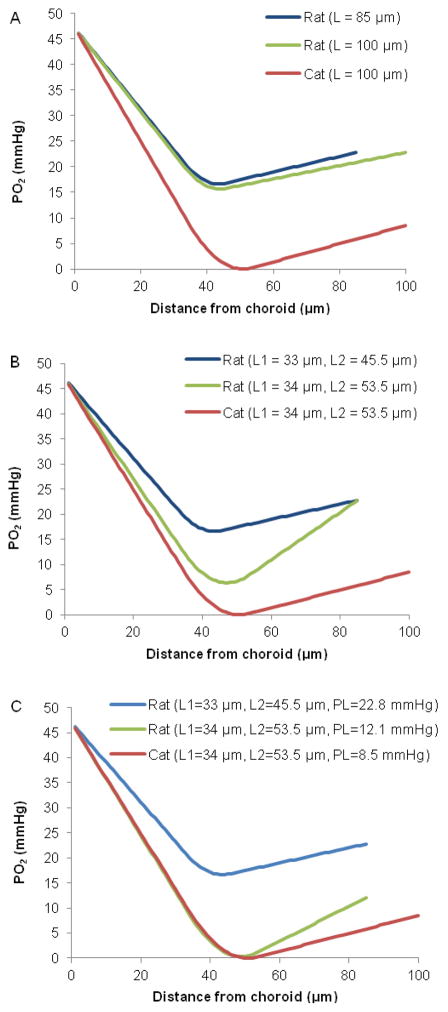
The purpose of this study was to investigate the oxygen distribution and consumption in the pigmented Long-Evans rat retina in vivo during dark and light adaptation, and to compare these results to previous work on cat and albino rat. Double-barreled microelectrodes recorded both intraretinal PO(2) depth profiles and the electroretinogram (ERG), which was used to identify the boundaries of the retina. Light adaptation decreased photoreceptor oxygen consumption per unit volume (Q(av)) from 3.0 ± 0.4 ml·100 g(-1) min(-1) (mean ± SEM) in darkness to 1.8 ± 0.2 ml·100 g(-1) min(-1) and increased minimum outer retinal PO(2) at the inner segments (P(min)) from 17.4 ± 3.0 to 29.9 ± 5.3 mmHg. The effects of light on outer retinal PO(2) and Q(av) were similar to those previously observed in cat, monkey, and albino rats; however, dark-adapted P(min) was higher in rat than cat. The parameters derived from fitting the oxygen diffusion model to the rat data were compared to those from cat. Oxygen consumption of the inner segments (Q(2)) and choroidal PO(2) (P(C)) in rat and cat were similar. P(min) was higher in rat than in cat for two reasons: first, rat photoreceptors have a shorter oxygen consuming region; and second, the retinal circulation supplied a greater fraction of consumed oxygen to rat photoreceptors. The average PO(2) across the inner retina (P(IR)) was not different in dark adaptation (25.4 ± 4.8 mmHg) and light adaptation (28.8 ± 5.4 mmHg) when measured from PO(2) profiles. However, with the microelectrode stationary at 9-18% retinal depth, a small consistent decrease in PO(2) occurred during illumination. Flickering light at 6 Hz decreased inner retinal PO(2) significantly more than an equivalent steady illumination, suggesting that changes in blood flow did not completely compensate for increased metabolism. This study comprehensively characterized rat retinal oxygenation in both light and dark, and determined the similarities and differences between rat and cat retinas.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Ahmed J, Braun RD, Dunn R, Jr, Linsenmeier RA. Oxygen distribution in the macaque retina. Invest Ophthalmol Vis Sci. 1993;34:516–521. - PubMed
-
- Alder VA, Yu DY, Cringle SJ. Vitreal oxygen tension measurements in the rat eye. Exp Eye Res. 1991;52:293–299. - PubMed
-
- Barnett V, Lewis T. Outliers in Statistical Data. John Wiley & Sons; New York: 1978.
-
- Bill A, Sperber GO. Aspects of oxygen and glucose consumption in the retina: effects of high intraocular pressure and light. Graefes Arch Clin Exp Ophthalmol. 1990;228:124–127. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
