

Nuclear ribosome biogenesis mediated by the DIM1A rRNA dimethylase is required for organized root growth and epidermal patterning in Arabidopsis
- PMID: 22829145
- PMCID: PMC3426118
- DOI: 10.1105/tpc.112.101022
Nuclear ribosome biogenesis mediated by the DIM1A rRNA dimethylase is required for organized root growth and epidermal patterning in Arabidopsis
Abstract
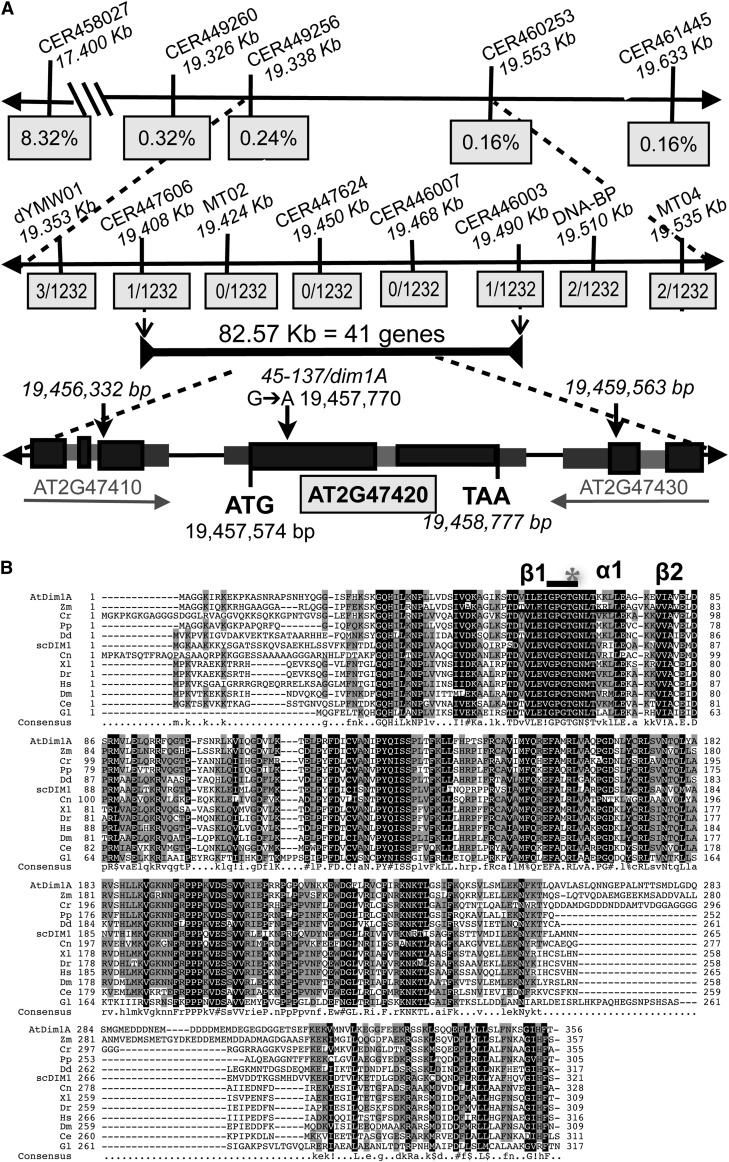
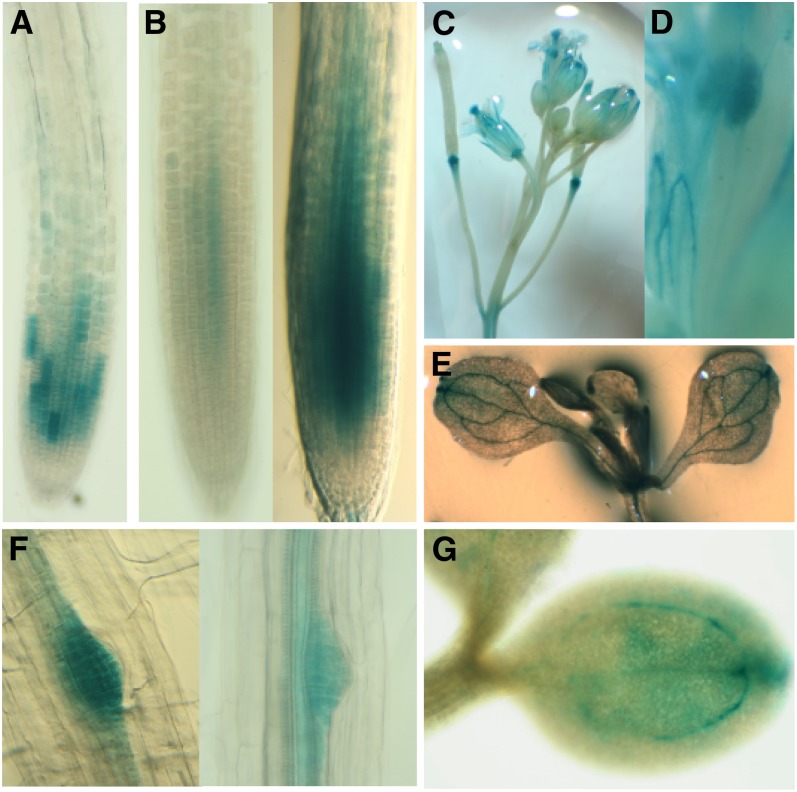
Position-dependent patterning of hair and non-hair cells in the Arabidopsis thaliana root epidermis is a powerful system to study the molecular basis of cell fate specification. Here, we report an epidermal patterning mutant affecting the ADENOSINE DIMETHYL TRANSFERASE 1A (DIM1A) rRNA dimethylase gene, predicted to participate in rRNA posttranscriptional processing and base modification. Consistent with a role in ribosome biogenesis, DIM1A is preferentially expressed in regions of rapid growth, and its product is nuclear localized with nucleolus enrichment. Furthermore, DIM1A preferentially accumulates in the developing hair cells, and the dim1A point mutant alters the cell-specific expression of the transcriptional regulators GLABRA2, CAPRICE, and WEREWOLF. Together, these findings suggest that establishment of cell-specific gene expression during root epidermis development is dependent upon proper ribosome biogenesis, possibly due to the sensitivity of the cell fate decision to relatively small differences in gene regulatory activities. Consistent with its effect on the predicted S-adenosyl-l-Met binding site, dim1A plants lack the two 18S rRNA base modifications but exhibit normal pre-rRNA processing. In addition to root epidermal defects, the dim1A mutant exhibits abnormal root meristem division, leaf development, and trichome branching. Together, these findings provide new insights into the importance of rRNA base modifications and translation regulation for plant growth and development.
Figures








Similar articles
-
Molecular Basis for a Cell Fate Switch in Response to Impaired Ribosome Biogenesis in the Arabidopsis Root Epidermis.Plant Cell. 2020 Jul;32(7):2402-2423. doi: 10.1105/tpc.19.00773. Epub 2020 May 5. Plant Cell. 2020. PMID: 32371546 Free PMC article.
-
A mutation of casein kinase 2 α4 subunit affects multiple developmental processes in Arabidopsis.Plant Cell Rep. 2016 May;35(5):1071-80. doi: 10.1007/s00299-016-1939-5. Epub 2016 Feb 16. Plant Cell Rep. 2016. PMID: 26883224
-
Positional signaling mediated by a receptor-like kinase in Arabidopsis.Science. 2005 Feb 18;307(5712):1111-3. doi: 10.1126/science.1105373. Epub 2004 Dec 23. Science. 2005. PMID: 15618487
-
The gene regulatory network for root epidermal cell-type pattern formation in Arabidopsis.J Exp Bot. 2009;60(5):1515-21. doi: 10.1093/jxb/ern339. Epub 2009 Jan 27. J Exp Bot. 2009. PMID: 19174459 Free PMC article. Review.
-
Computational modeling of epidermal cell fate determination systems.Curr Opin Plant Biol. 2013 Feb;16(1):5-10. doi: 10.1016/j.pbi.2012.12.003. Epub 2013 Jan 1. Curr Opin Plant Biol. 2013. PMID: 23287386 Review.
Cited by
-
An epigenetic basis of inbreeding depression in maize.Sci Adv. 2021 Aug 27;7(35):eabg5442. doi: 10.1126/sciadv.abg5442. Print 2021 Aug. Sci Adv. 2021. PMID: 34452913 Free PMC article.
-
Molecular Basis for a Cell Fate Switch in Response to Impaired Ribosome Biogenesis in the Arabidopsis Root Epidermis.Plant Cell. 2020 Jul;32(7):2402-2423. doi: 10.1105/tpc.19.00773. Epub 2020 May 5. Plant Cell. 2020. PMID: 32371546 Free PMC article.
-
Structural and catalytic roles of the human 18S rRNA methyltransferases DIMT1 in ribosome assembly and translation.J Biol Chem. 2020 Aug 21;295(34):12058-12070. doi: 10.1074/jbc.RA120.014236. Epub 2020 Jul 2. J Biol Chem. 2020. PMID: 32616653 Free PMC article.
-
Two Nucleolar Proteins, GDP1 and OLI2, Function As Ribosome Biogenesis Factors and Are Preferentially Involved in Promotion of Leaf Cell Proliferation without Strongly Affecting Leaf Adaxial-Abaxial Patterning in Arabidopsis thaliana.Front Plant Sci. 2018 Jan 9;8:2240. doi: 10.3389/fpls.2017.02240. eCollection 2017. Front Plant Sci. 2018. PMID: 29375609 Free PMC article.
-
Histone Deacetylases HD2A and HD2B Undergo Feedback Regulation by ABA and Modulate Drought Tolerance via Mediating ABA-Induced Transcriptional Repression.Genes (Basel). 2023 May 30;14(6):1199. doi: 10.3390/genes14061199. Genes (Basel). 2023. PMID: 37372378 Free PMC article.
References
-
- Abbasi N., Kim H.B., Park N.-I., Kim H.-S., Kim Y.-K., Park Y.-I., Choi S.-B. (2010). APUM23, a nucleolar Puf domain protein, is involved in pre-ribosomal RNA processing and normal growth patterning in Arabidopsis. Plant J. 64: 960–976 - PubMed
-
- Berger F., Hung C.Y., Dolan L., Schiefelbein J. (1998). Control of cell division in the root epidermis of Arabidopsis thaliana. Dev. Biol. 194: 235–245 - PubMed
-
- Bernhardt C., Lee M.M., Gonzalez A., Zhang F., Lloyd A., Schiefelbein J. (2003). The bHLH genes GLABRA3 (GL3) and ENHANCER OF GLABRA3 (EGL3) specify epidermal cell fate in the Arabidopsis root. Development 130: 6431–6439 - PubMed
-
- Bernhardt C., Zhao M., Gonzalez A., Lloyd A., Schiefelbein J. (2005). The bHLH genes GL3 and EGL3 participate in an intercellular regulatory circuit that controls cell patterning in the Arabidopsis root epidermis. Development 132: 291–298 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
