

Structural dynamics of nucleosomes at single-molecule resolution
- PMID: 22831768
- PMCID: PMC3669752
- DOI: 10.1016/j.tibs.2012.06.006
Structural dynamics of nucleosomes at single-molecule resolution
Abstract
The detailed mechanisms of how DNA that is assembled around a histone core can be accessed by DNA-binding proteins for transcription, replication, or repair, remain elusive nearly 40 years after Kornberg's nucleosome model was proposed. Uncovering the structural dynamics of nucleosomes is a crucial step in elucidating the mechanisms regulating genome accessibility. This requires the deconvolution of multiple structural states within an ensemble. Recent advances in single-molecule methods enable unprecedented efficiency in examining subpopulation dynamics. In this review, we summarize studies of nucleosome structure and dynamics from single-molecule approaches and how they advance our understanding of the mechanisms that govern DNA transactions.
Published by Elsevier Ltd.
Figures
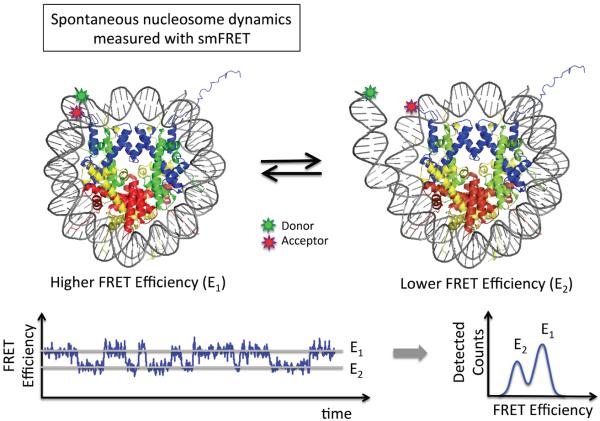
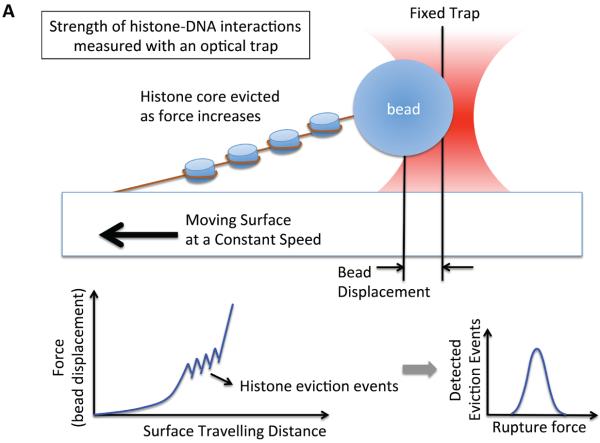
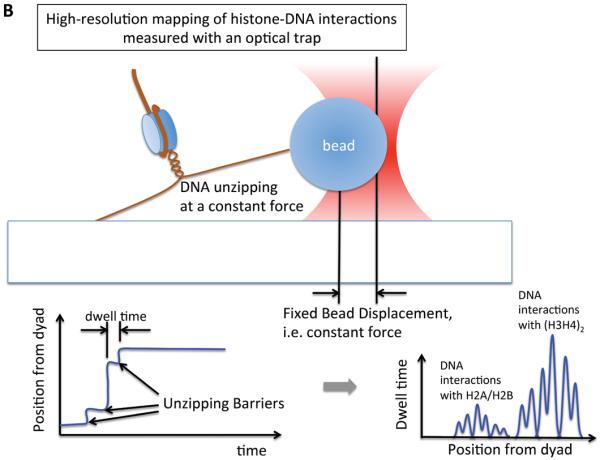
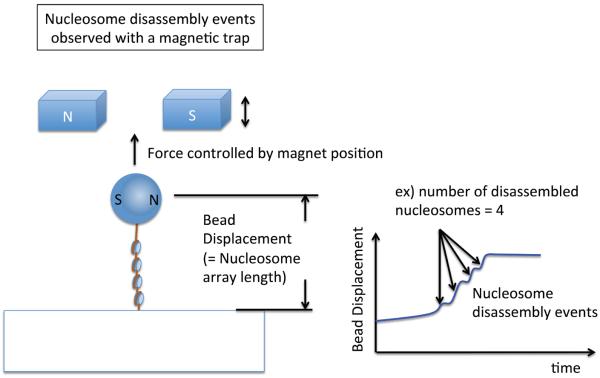
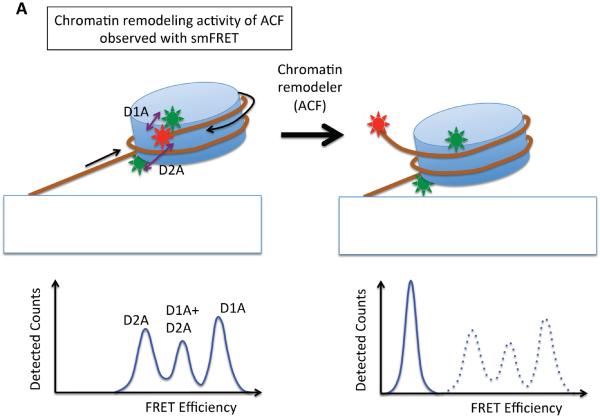
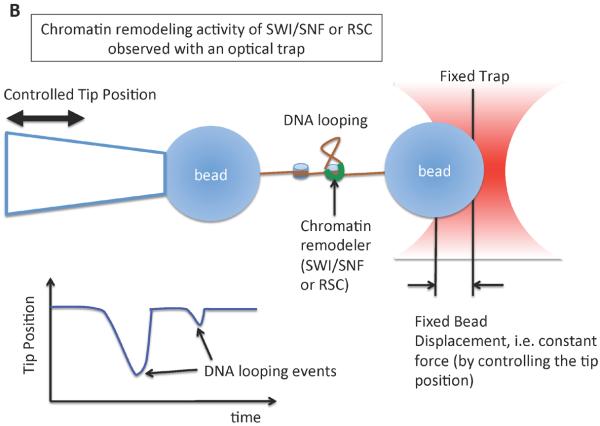
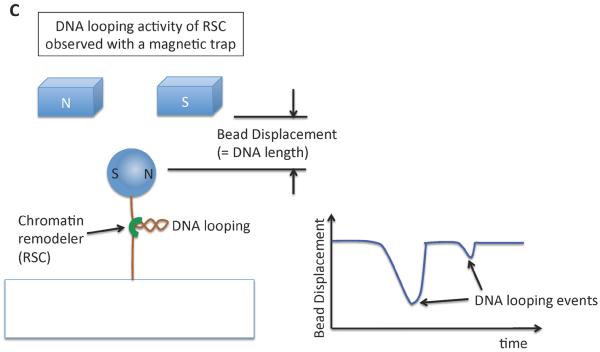
References
-
- Kornberg RD. Chromatin structure: a repeating unit of histones and DNA. Science. 1974;184:868–871. - PubMed
-
- Kornberg RD, Thomas JO. Chromatin structure; oligomers of the histones. Science. 1974;184:865–868. - PubMed
-
- Kornberg RD. Structure of chromatin. Annu Rev Biochem. 1977;46:931–954. - PubMed
-
- Richmond TJ, et al. Structure of the nucleosome core particle at 7 A resolution. Nature. 1984;311:532–537. - PubMed
-
- Klug A, et al. A low resolution structure for the histone core of the nucleosome. Nature. 1980;287:509–516. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
