

Crystal structure of a minimal eIF4E-Cup complex reveals a general mechanism of eIF4E regulation in translational repression
- PMID: 22832024
- PMCID: PMC3425778
- DOI: 10.1261/rna.033639.112
Crystal structure of a minimal eIF4E-Cup complex reveals a general mechanism of eIF4E regulation in translational repression
Abstract
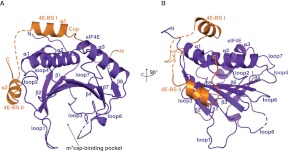
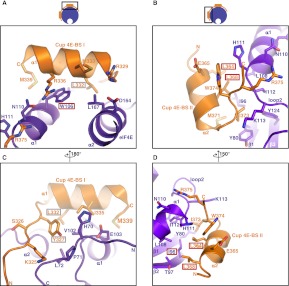
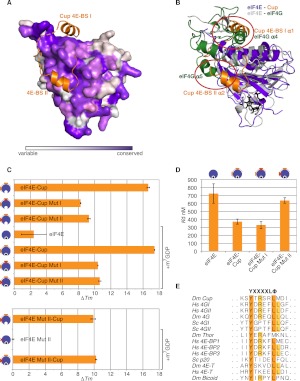
Cup is an eIF4E-binding protein (4E-BP) that plays a central role in translational regulation of localized mRNAs during early Drosophila development. In particular, Cup is required for repressing translation of the maternally contributed oskar, nanos, and gurken mRNAs, all of which are essential for embryonic body axis determination. Here, we present the 2.8 Å resolution crystal structure of a minimal eIF4E-Cup assembly, consisting of the interacting regions of the two proteins. In the structure, two separate segments of Cup contact two orthogonal faces of eIF4E. The eIF4E-binding consensus motif of Cup (YXXXXLΦ) binds the convex side of eIF4E similarly to the consensus of other eIF4E-binding proteins, such as 4E-BPs and eIF4G. The second, noncanonical, eIF4E-binding site of Cup binds laterally and perpendicularly to the eIF4E β-sheet. Mutations of Cup at this binding site were shown to reduce binding to eIF4E and to promote the destabilization of the associated mRNA. Comparison with the binding mode of eIF4G to eIF4E suggests that Cup and eIF4G binding would be mutually exclusive at both binding sites. This shows how a common molecular surface of eIF4E might recognize different proteins acting at different times in the same pathway. The structure provides insight into the mechanism by which Cup disrupts eIF4E-eIF4G interaction and has broader implications for understanding the role of 4E-BPs in translational regulation.
Figures

Similar articles
-
4E-BPs require non-canonical 4E-binding motifs and a lateral surface of eIF4E to repress translation.Nat Commun. 2014 Sep 2;5:4790. doi: 10.1038/ncomms5790. Nat Commun. 2014. PMID: 25179781 Free PMC article.
-
The Structures of eIF4E-eIF4G Complexes Reveal an Extended Interface to Regulate Translation Initiation.Mol Cell. 2016 Nov 3;64(3):467-479. doi: 10.1016/j.molcel.2016.09.020. Epub 2016 Oct 20. Mol Cell. 2016. PMID: 27773676
-
Drosophila Cup is an eIF4E-binding protein that functions in Smaug-mediated translational repression.EMBO J. 2004 Jan 14;23(1):150-9. doi: 10.1038/sj.emboj.7600026. Epub 2003 Dec 11. EMBO J. 2004. PMID: 14685270 Free PMC article.
-
Control of the eIF4E activity: structural insights and pharmacological implications.Cell Mol Life Sci. 2021 Nov;78(21-22):6869-6885. doi: 10.1007/s00018-021-03938-z. Epub 2021 Sep 19. Cell Mol Life Sci. 2021. PMID: 34541613 Free PMC article. Review.
-
Contrasting mechanisms of regulating translation of specific Drosophila germline mRNAs at the level of 5'-cap structure binding.Biochem Soc Trans. 2005 Dec;33(Pt 6):1544-6. doi: 10.1042/BST0331544. Biochem Soc Trans. 2005. PMID: 16246166 Review.
Cited by
-
Structural basis for LeishIF4E-1 modulation by an interacting protein in the human parasite Leishmania major.Nucleic Acids Res. 2018 Apr 20;46(7):3791-3801. doi: 10.1093/nar/gky194. Nucleic Acids Res. 2018. PMID: 29562352 Free PMC article.
-
mRNA localization in metazoans: A structural perspective.RNA Biol. 2017 Nov 2;14(11):1473-1484. doi: 10.1080/15476286.2017.1338231. Epub 2017 Jul 31. RNA Biol. 2017. PMID: 28640665 Free PMC article. Review.
-
Analysis of the interacting partners eIF4F and 3'-CITE required for Melon necrotic spot virus cap-independent translation.Mol Plant Pathol. 2017 Jun;18(5):635-648. doi: 10.1111/mpp.12422. Epub 2016 Jul 27. Mol Plant Pathol. 2017. PMID: 27145354 Free PMC article.
-
The crystal structure of the PB2 cap-binding domain of influenza B virus reveals a novel cap recognition mechanism.J Biol Chem. 2015 Apr 3;290(14):9141-9. doi: 10.1074/jbc.M115.636464. Epub 2015 Feb 17. J Biol Chem. 2015. PMID: 25691568 Free PMC article.
-
Investigating the consequences of eIF4E2 (4EHP) interaction with 4E-transporter on its cellular distribution in HeLa cells.PLoS One. 2013 Aug 21;8(8):e72761. doi: 10.1371/journal.pone.0072761. eCollection 2013. PLoS One. 2013. PMID: 23991149 Free PMC article.
References
-
- Besse F, Ephrussi A 2008. Translational control of localized mRNAs: Restricting protein synthesis in space and time. Nat Rev Mol Cell Biol 9: 971–980 - PubMed
-
- Brown CJ, McNae I, Fischer PM, Walkinshaw MD 2007. Crystallographic and mass spectrometric characterisation of eIF4E with N7-alkylated cap derivatives. J Mol Biol 372: 7–15 - PubMed
-
- Brown CJ, Verma CS, Walkinshaw MD, Lane DP 2009. Crystallization of eIF4E complexed with eIF4GI peptide and glycerol reveals distinct structural differences around the cap-binding site. Cell Cycle 8: 1905–1911 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases