

Fgfr-Ras-MAPK signaling is required for apical constriction via apical positioning of Rho-associated kinase during mechanosensory organ formation
- PMID: 22833124
- PMCID: PMC3413159
- DOI: 10.1242/dev.082271
Fgfr-Ras-MAPK signaling is required for apical constriction via apical positioning of Rho-associated kinase during mechanosensory organ formation
Erratum in
- Development. 2012 Sep 15;139(18):3467
Abstract
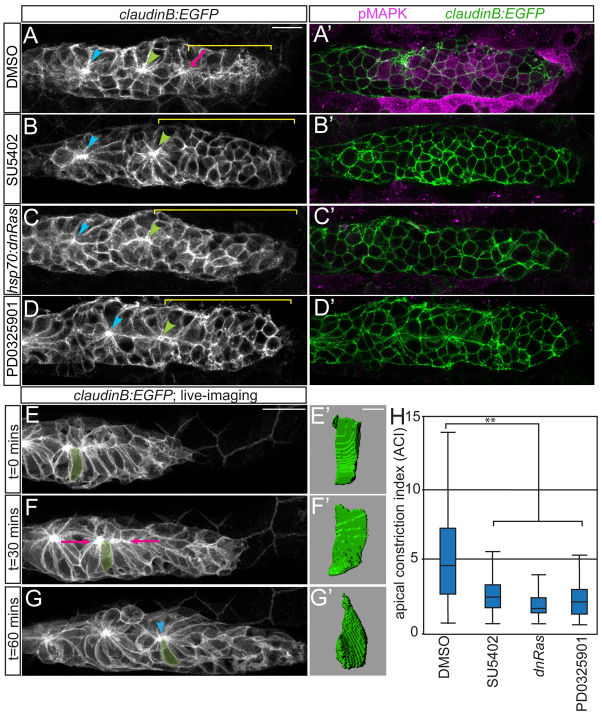
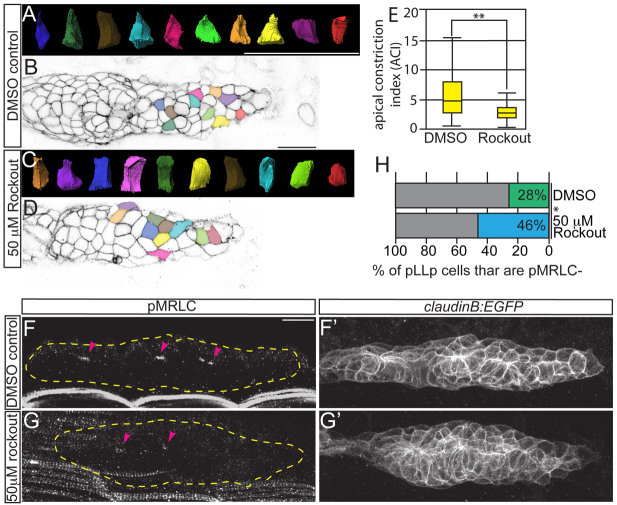
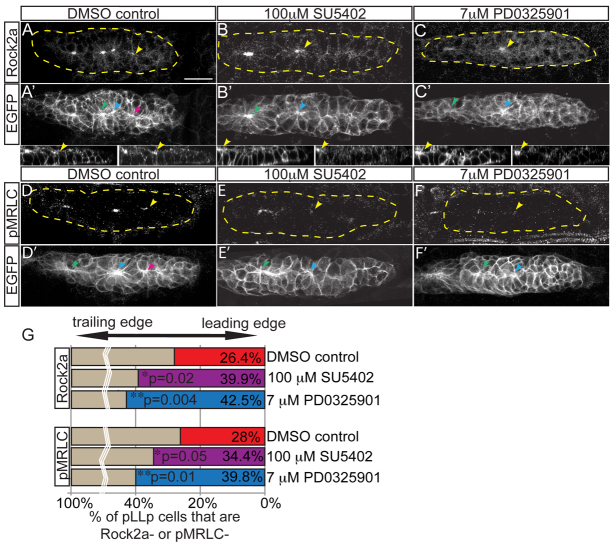
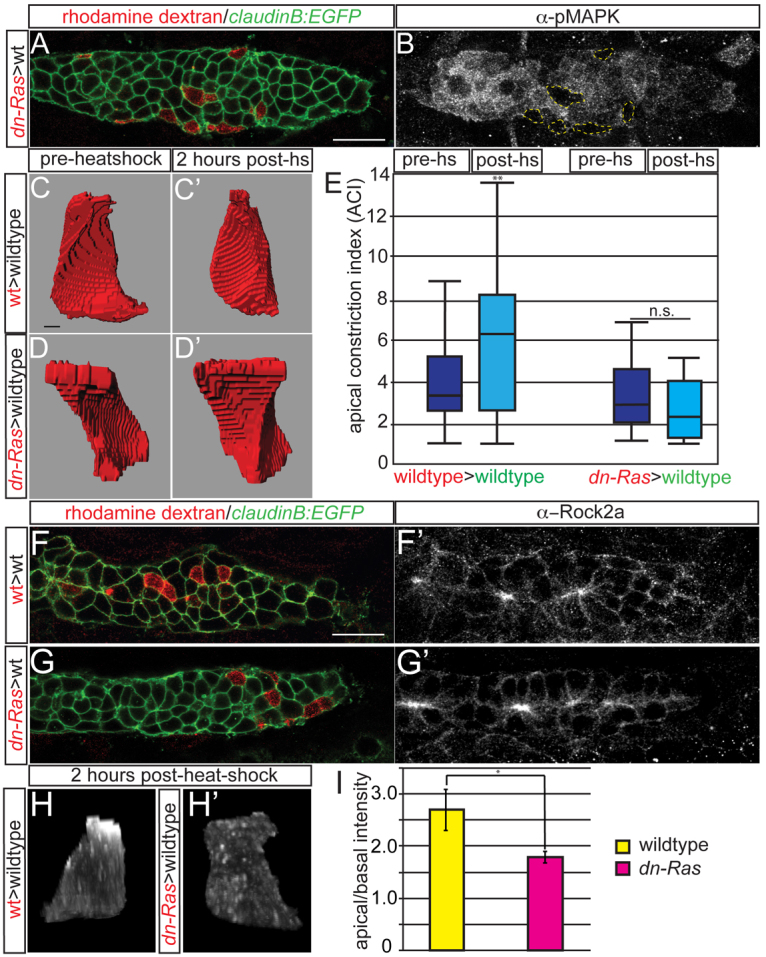
Many morphogenetic movements during development require the formation of transient intermediates called rosettes. Within rosettes, cells are polarized with apical ends constricted towards the rosette center and nuclei basally displaced. Whereas the polarity and cytoskeletal machinery establishing these structures has been extensively studied, the extracellular cues and intracellular signaling cascades that promote their formation are not well understood. We examined how extracellular Fibroblast growth factor (Fgf) signals regulate rosette formation in the zebrafish posterior lateral line primordium (pLLp), a group of ∼100 cells that migrates along the trunk during embryonic development to form the lateral line mechanosensory system. During migration, the pLLp deposits rosettes from the trailing edge, while cells are polarized and incorporated into nascent rosettes in the leading region. Fgf signaling was previously shown to be crucial for rosette formation in the pLLp. We demonstrate that activation of Fgf receptor (Fgfr) induces intracellular Ras-MAPK, which is required for apical constriction and rosette formation in the pLLp. Inhibiting Fgfr-Ras-MAPK leads to loss of apically localized Rho-associated kinase (Rock) 2a, which results in failed actomyosin cytoskeleton activation. Using mosaic analyses, we show that a cell-autonomous Ras-MAPK signal is required for apical constriction and Rock2a localization. We propose a model whereby activated Fgfr signals through Ras-MAPK to induce apical localization of Rock2a in a cell-autonomous manner, activating the actomyosin network to promote apical constriction and rosette formation in the pLLp. This mechanism presents a novel cellular strategy for driving cell shape changes.
Figures
Similar articles
-
RhoA GEF Mcf2lb regulates rosette integrity during collective cell migration.Development. 2024 Jan 1;151(1):dev201898. doi: 10.1242/dev.201898. Epub 2024 Jan 2. Development. 2024. PMID: 38165177 Free PMC article.
-
Shroom3 is required downstream of FGF signalling to mediate proneuromast assembly in zebrafish.Development. 2012 Dec;139(24):4571-81. doi: 10.1242/dev.083253. Epub 2012 Nov 7. Development. 2012. PMID: 23136387
-
RhoA GEF Mcf2lb regulates rosette integrity during collective cell migration.bioRxiv [Preprint]. 2023 Apr 27:2023.04.19.537573. doi: 10.1101/2023.04.19.537573. bioRxiv. 2023. Update in: Development. 2024 Jan 1;151(1):dev201898. doi: 10.1242/dev.201898. PMID: 37131612 Free PMC article. Updated. Preprint.
-
Signaling pathways regulating zebrafish lateral line development.Curr Biol. 2009 May 12;19(9):R381-6. doi: 10.1016/j.cub.2009.03.057. Curr Biol. 2009. PMID: 19439264 Review.
-
The roles and regulation of multicellular rosette structures during morphogenesis.Development. 2014 Jul;141(13):2549-58. doi: 10.1242/dev.101444. Development. 2014. PMID: 24961796 Free PMC article. Review.
Cited by
-
Prognostic value of FGFR1 expression and amplification in patients with HNSCC: A systematic review and meta-analysis.PLoS One. 2021 May 14;16(5):e0251202. doi: 10.1371/journal.pone.0251202. eCollection 2021. PLoS One. 2021. PMID: 33989301 Free PMC article.
-
Gβ1 controls collective cell migration by regulating the protrusive activity of leader cells in the posterior lateral line primordium.Dev Biol. 2014 Jan 15;385(2):316-27. doi: 10.1016/j.ydbio.2013.10.027. Epub 2013 Nov 4. Dev Biol. 2014. PMID: 24201188 Free PMC article.
-
Amotl2a interacts with the Hippo effector Yap1 and the Wnt/β-catenin effector Lef1 to control tissue size in zebrafish.Elife. 2015 Sep 3;4:e08201. doi: 10.7554/eLife.08201. Elife. 2015. PMID: 26335201 Free PMC article.
-
The molecular mechanisms underlying lens fiber elongation.Exp Eye Res. 2017 Mar;156:41-49. doi: 10.1016/j.exer.2016.03.016. Epub 2016 Mar 23. Exp Eye Res. 2017. PMID: 27015931 Free PMC article. Review.
-
Pulses of RhoA signaling stimulate actin polymerization and flow in protrusions to drive collective cell migration.Curr Biol. 2024 Jan 22;34(2):245-259.e8. doi: 10.1016/j.cub.2023.11.044. Epub 2023 Dec 13. Curr Biol. 2024. PMID: 38096821 Free PMC article.
References
-
- Abramoff M. D. (2004). Image processing with ImageJ. Biophotonics International 11, 36–42
-
- Andermann P., Ungos J., Raible D. W. (2002). Neurogenin1 defines zebrafish cranial sensory ganglia precursors. Dev. Biol. 251, 45–58 - PubMed
-
- Ghysen A., Dambly-Chaudière C. (2007). The lateral line microcosmos. Genes Dev. 21, 2118–2130 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
