

Functions of S100 proteins
Abstract
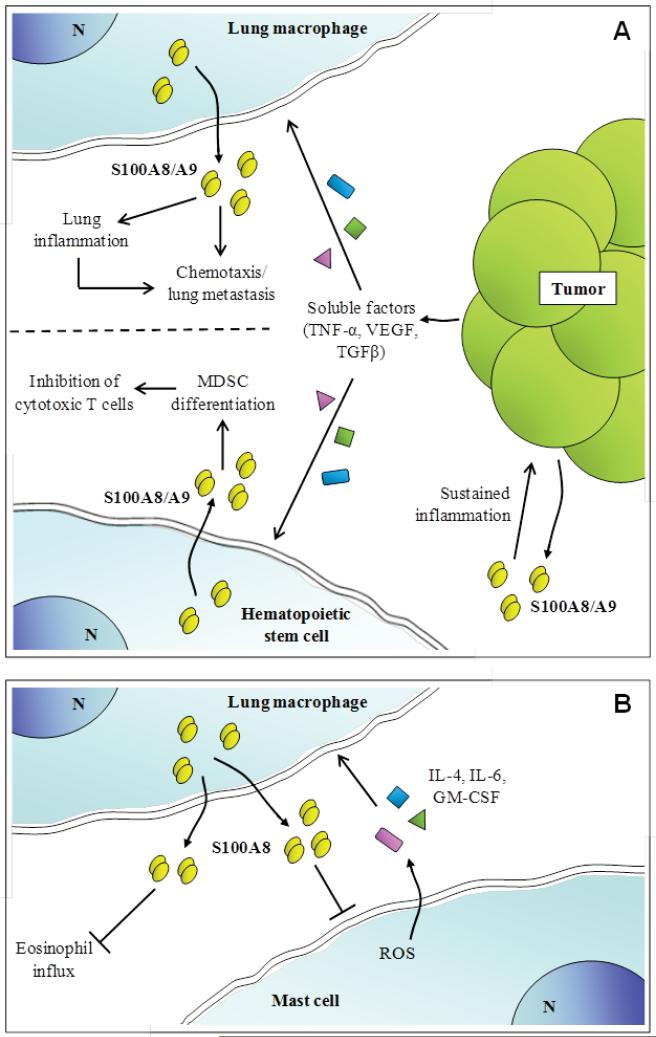
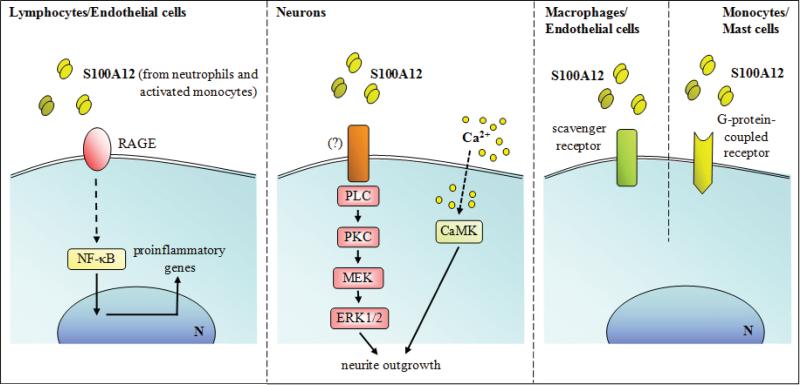
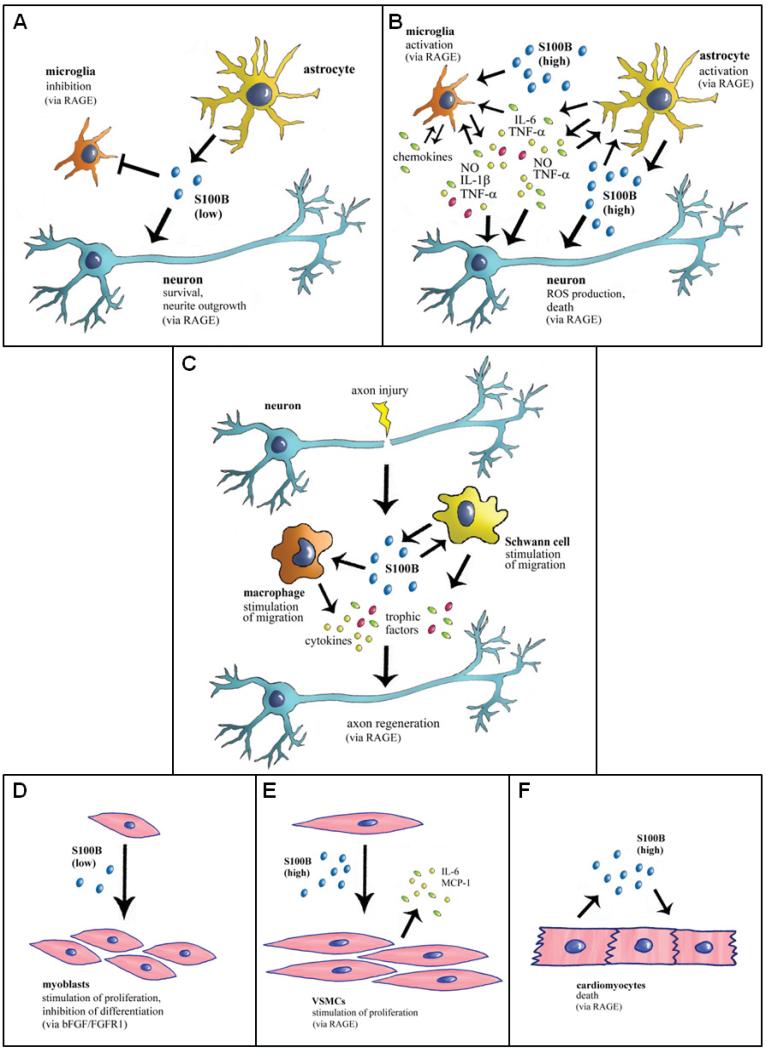
The S100 protein family consists of 24 members functionally distributed into three main subgroups: those that only exert intracellular regulatory effects, those with intracellular and extracellular functions and those which mainly exert extracellular regulatory effects. S100 proteins are only expressed in vertebrates and show cell-specific expression patterns. In some instances, a particular S100 protein can be induced in pathological circumstances in a cell type that does not express it in normal physiological conditions. Within cells, S100 proteins are involved in aspects of regulation of proliferation, differentiation, apoptosis, Ca2+ homeostasis, energy metabolism, inflammation and migration/invasion through interactions with a variety of target proteins including enzymes, cytoskeletal subunits, receptors, transcription factors and nucleic acids. Some S100 proteins are secreted or released and regulate cell functions in an autocrine and paracrine manner via activation of surface receptors (e.g. the receptor for advanced glycation end-products and toll-like receptor 4), G-protein-coupled receptors, scavenger receptors, or heparan sulfate proteoglycans and N-glycans. Extracellular S100A4 and S100B also interact with epidermal growth factor and basic fibroblast growth factor, respectively, thereby enhancing the activity of the corresponding receptors. Thus, extracellular S100 proteins exert regulatory activities on monocytes/macrophages/microglia, neutrophils, lymphocytes, mast cells, articular chondrocytes, endothelial and vascular smooth muscle cells, neurons, astrocytes, Schwann cells, epithelial cells, myoblasts and cardiomyocytes, thereby participating in innate and adaptive immune responses, cell migration and chemotaxis, tissue development and repair, and leukocyte and tumor cell invasion.
Figures
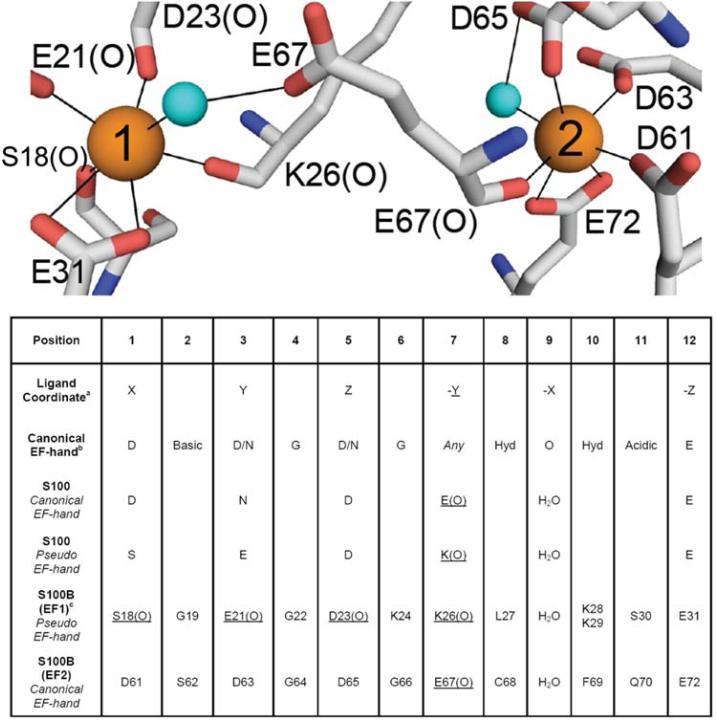
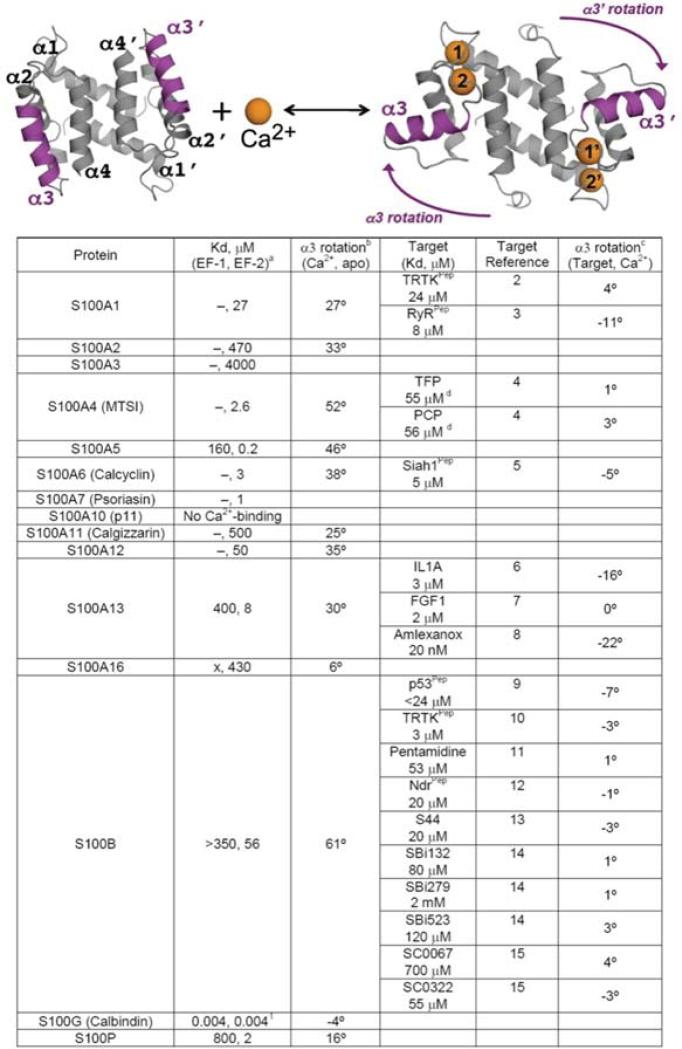
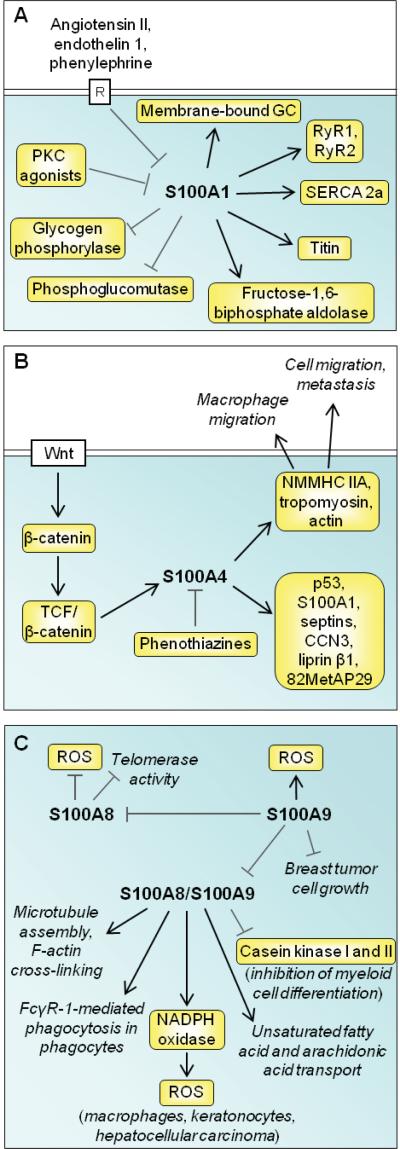
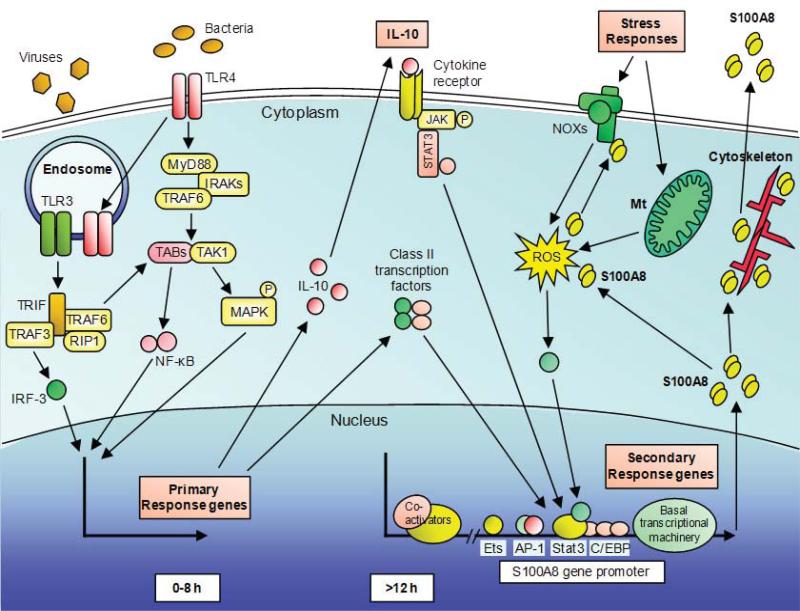
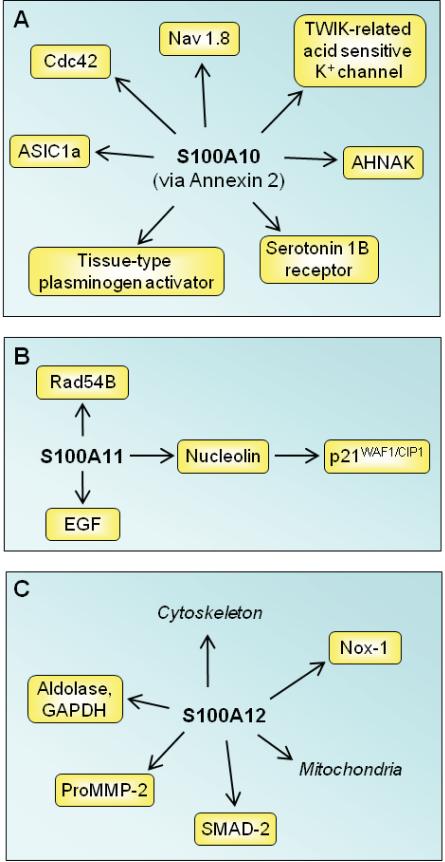
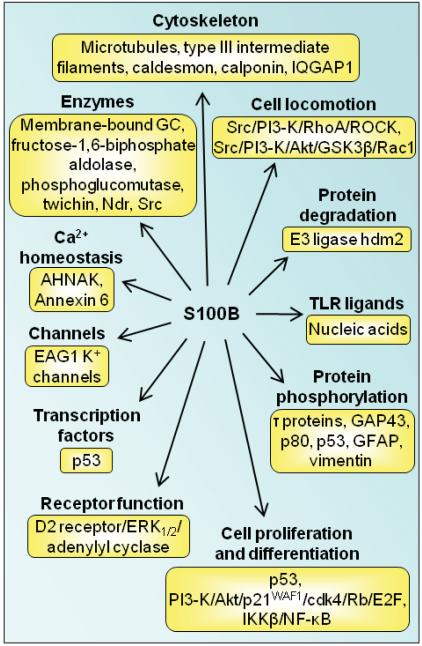
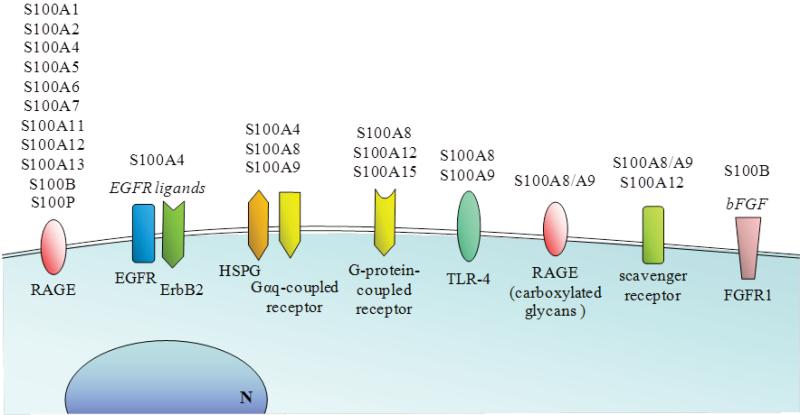
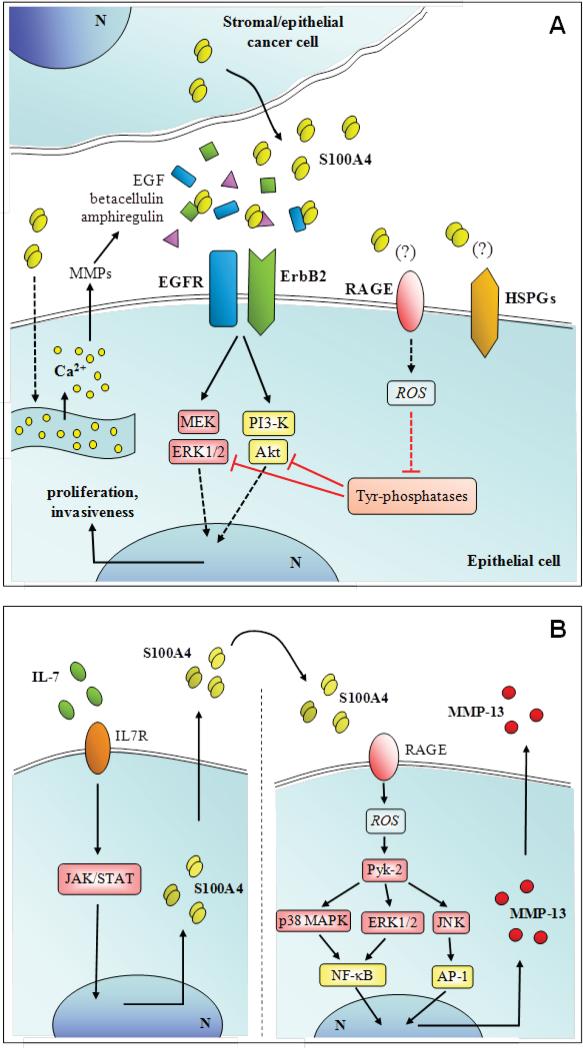
References
-
- Carafoli E, Klee C. Calcium as a Cellular Regulator. Oxford University Press; New York: 1999.
-
- Permyakov EA, Kretsinger RH. Cell signaling, beyond cytosolic calcium in eukaryotes. J Inorg Biochem. 2009;103:77–86. - PubMed
-
- Donato R. S100: a multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int J Biochem Cell Biol. 2001;2001;33:637–8. - PubMed
-
- Marenholz I, Heizmann CW, Fritz G. S100 proteins in mouse and man: from evolution to function and pathology (including an update of the nomenclature). Biochem Biophys Res Commun. 2004;322:1111–22. - PubMed
-
- Donato R. RAGE: a single receptor for several ligands and different cellular responses: the case of certain S100 proteins. Curr Mol Med. 2007;7:711–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous