

Drug experience epigenetically primes Fosb gene inducibility in rat nucleus accumbens
- PMID: 22836260
- PMCID: PMC3417057
- DOI: 10.1523/JNEUROSCI.1290-12.2012
Drug experience epigenetically primes Fosb gene inducibility in rat nucleus accumbens
Abstract
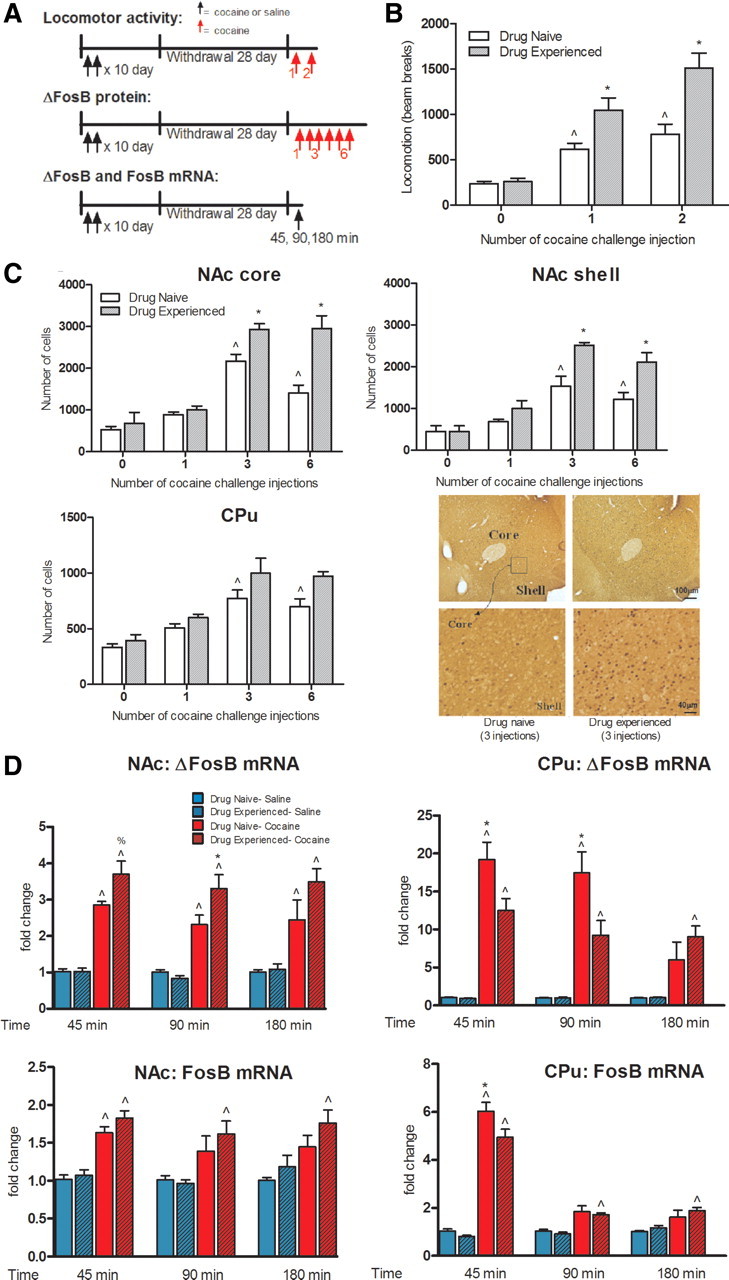
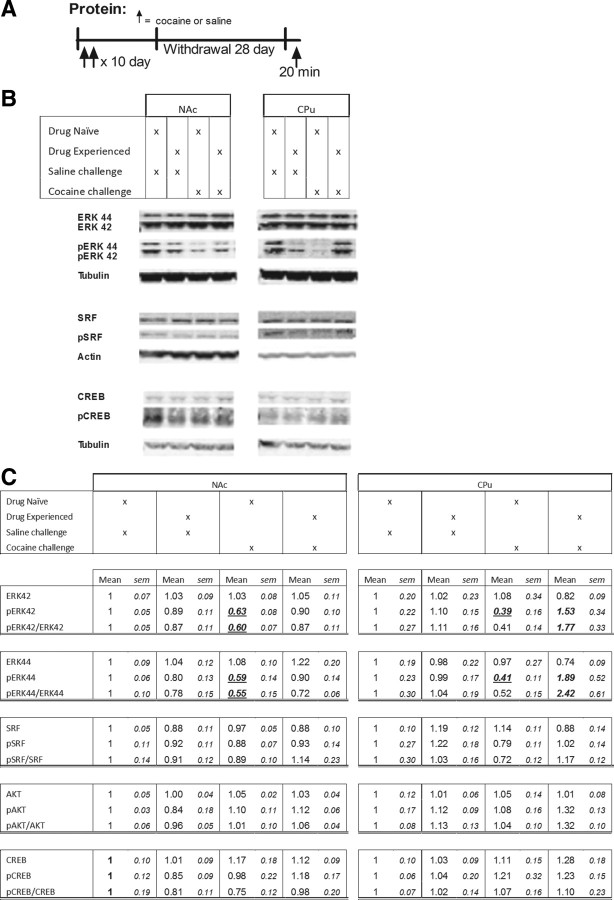
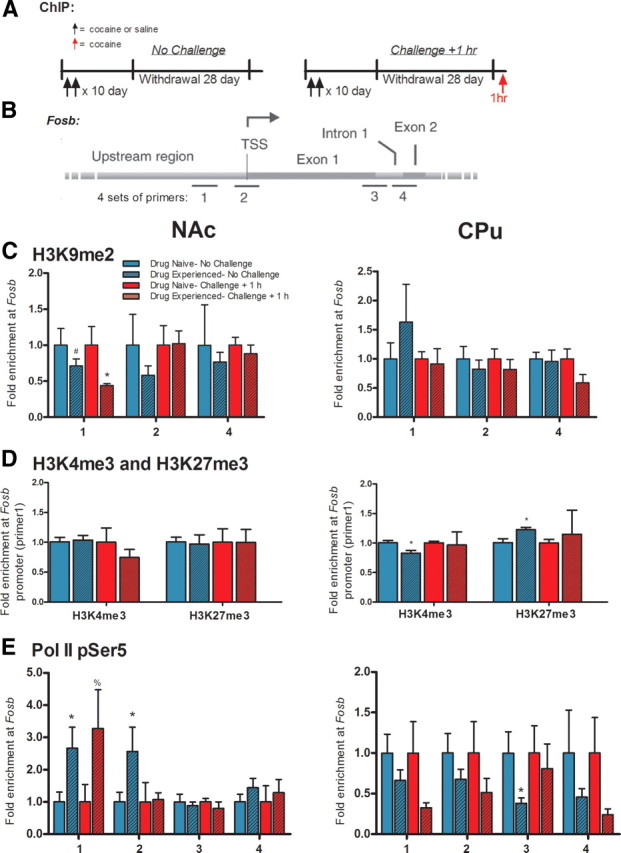
ΔFosB, a Fosb gene product, is induced in nucleus accumbens (NAc) and caudate-putamen (CPu) by repeated exposure to drugs of abuse such as cocaine. This induction contributes to aberrant patterns of gene expression and behavioral abnormalities seen with repeated drug exposure. Here, we assessed whether a remote history of cocaine exposure in rats might alter inducibility of the Fosb gene elicited by subsequent drug exposure. We show that prior chronic cocaine administration, followed by extended withdrawal, increases inducibility of Fosb in NAc, as evidenced by greater acute induction of ΔFosB mRNA and faster accumulation of ΔFosB protein after repeated cocaine reexposure. No such primed Fosb induction was observed in CPu; in fact, subsequent acute induction of ΔFosB mRNA was suppressed in CPu. These abnormal patterns of Fosb expression are associated with chromatin modifications at the Fosb gene promoter. Prior chronic cocaine administration induces a long-lasting increase in RNA polymerase II (Pol II) binding at the Fosb promoter in NAc only, suggesting that Pol II "stalling" primes Fosb for induction in this region upon reexposure to cocaine. A cocaine challenge then triggers the release of Pol II from the gene promoter, allowing for more rapid Fosb transcription. A cocaine challenge also decreases repressive histone modifications at the Fosb promoter in NAc, but increases such repressive marks and decreases activating marks in CPu. These results provide new insight into the chromatin dynamics at the Fosb promoter and reveal a novel mechanism for primed Fosb induction in NAc upon reexposure to cocaine.
Figures
Similar articles
-
Fosb Induction in Nucleus Accumbens by Cocaine Is Regulated by E2F3a.eNeuro. 2019 Apr 1;6(2):ENEURO.0325-18.2019. doi: 10.1523/ENEURO.0325-18.2019. eCollection 2019 Mar-Apr. eNeuro. 2019. PMID: 30963104 Free PMC article.
-
Serum response factor and cAMP response element binding protein are both required for cocaine induction of ΔFosB.J Neurosci. 2012 May 30;32(22):7577-84. doi: 10.1523/JNEUROSCI.1381-12.2012. J Neurosci. 2012. PMID: 22649236 Free PMC article.
-
Striatal regulation of ΔFosB, FosB, and cFos during cocaine self-administration and withdrawal.J Neurochem. 2010 Oct;115(1):112-22. doi: 10.1111/j.1471-4159.2010.06907.x. Epub 2010 Aug 3. J Neurochem. 2010. PMID: 20633205 Free PMC article.
-
Comparison of cocaine- and methamphetamine-evoked dopamine and glutamate overflow in somatodendritic and terminal field regions of the rat brain during acute, chronic, and early withdrawal conditions.Ann N Y Acad Sci. 2001 Jun;937:93-120. doi: 10.1111/j.1749-6632.2001.tb03560.x. Ann N Y Acad Sci. 2001. PMID: 11458542 Review.
-
∆FosB: a transcriptional regulator of stress and antidepressant responses.Eur J Pharmacol. 2015 Apr 15;753:66-72. doi: 10.1016/j.ejphar.2014.10.034. Epub 2014 Nov 7. Eur J Pharmacol. 2015. PMID: 25446562 Free PMC article. Review.
Cited by
-
Epigenetic Effects of Addictive Drugs in the Nucleus Accumbens.Front Mol Neurosci. 2022 Jun 23;15:828055. doi: 10.3389/fnmol.2022.828055. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35813068 Free PMC article. Review.
-
Histone arginine methylation in cocaine action in the nucleus accumbens.Proc Natl Acad Sci U S A. 2016 Aug 23;113(34):9623-8. doi: 10.1073/pnas.1605045113. Epub 2016 Aug 9. Proc Natl Acad Sci U S A. 2016. PMID: 27506785 Free PMC article.
-
Cocaine Self-administration Alters Transcriptome-wide Responses in the Brain's Reward Circuitry.Biol Psychiatry. 2018 Dec 15;84(12):867-880. doi: 10.1016/j.biopsych.2018.04.009. Epub 2018 Apr 25. Biol Psychiatry. 2018. PMID: 29861096 Free PMC article.
-
Psychostimulant-Induced Adaptations in Nucleus Accumbens Glutamatergic Transmission.Cold Spring Harb Perspect Med. 2020 Dec 1;10(12):a039255. doi: 10.1101/cshperspect.a039255. Cold Spring Harb Perspect Med. 2020. PMID: 31964644 Free PMC article. Review.
-
Current perspectives on the neurobiology of drug addiction: a focus on genetics and factors regulating gene expression.ISRN Neurol. 2012;2012:972607. doi: 10.5402/2012/972607. Epub 2012 Oct 14. ISRN Neurol. 2012. PMID: 23097719 Free PMC article.
References
-
- Bataille AR, Jeronimo C, Jacques PÉ, Laramée L, Fortin MÈ, Forest A, Bergeron M, Hanes SD, Robert F. A universal RNA polymerase II CTD cycle is orchestrated by complex interplays between kinase, phosphatase, and isomerase enzymes along genes. Mol Cell. 2012;45:158–170. - PubMed
-
- Boudreau AC, Reimers JM, Milovanovic M, Wolf ME. Cell surface AMPA receptors in the rat nucleus accumbens increase during cocaine withdrawal but internalize after cocaine challenge in association with altered activation of mitogen-activated protein kinases. J Neurosci. 2007;27:10621–10635. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous