

TLR7 recognition is dispensable for influenza virus A infection but important for the induction of hemagglutinin-specific antibodies in response to the 2009 pandemic split vaccine in mice
- PMID: 22837197
- PMCID: PMC3457183
- DOI: 10.1128/JVI.01064-12
TLR7 recognition is dispensable for influenza virus A infection but important for the induction of hemagglutinin-specific antibodies in response to the 2009 pandemic split vaccine in mice
Abstract
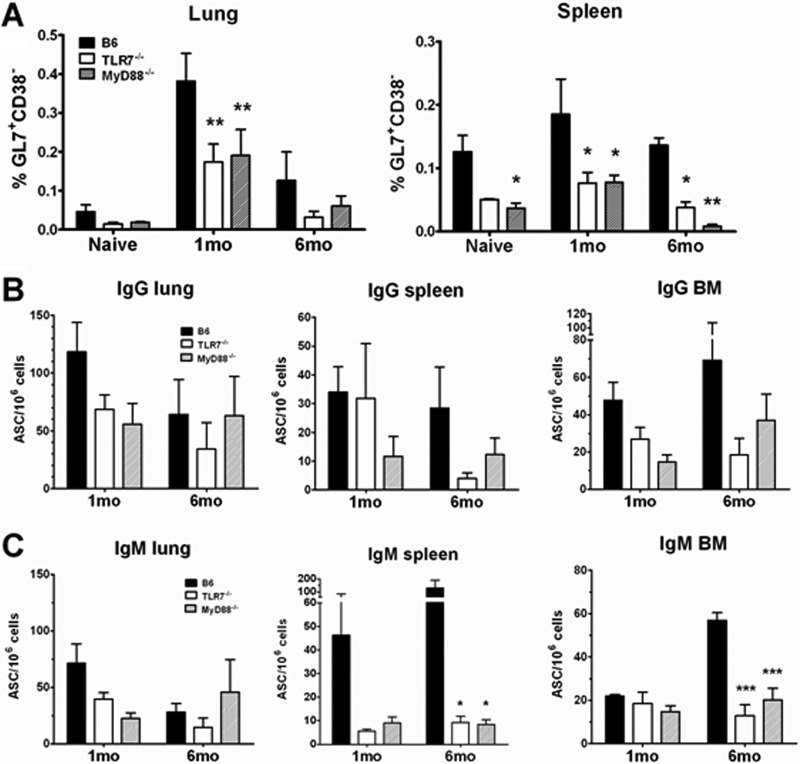
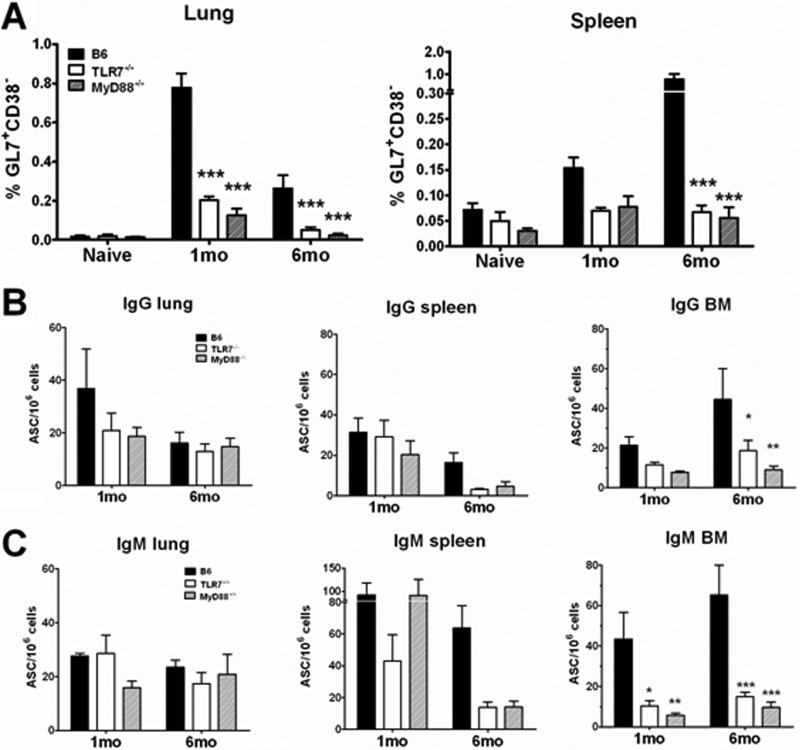
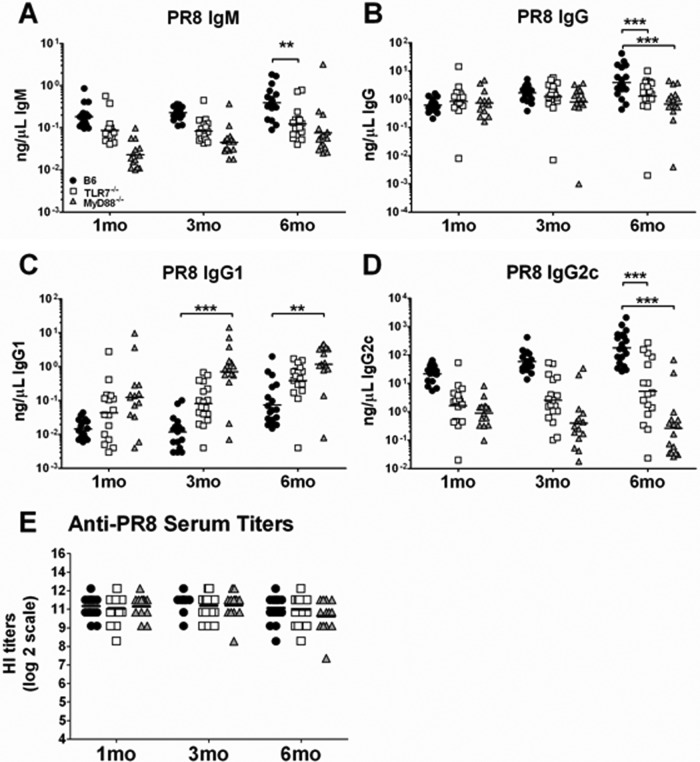
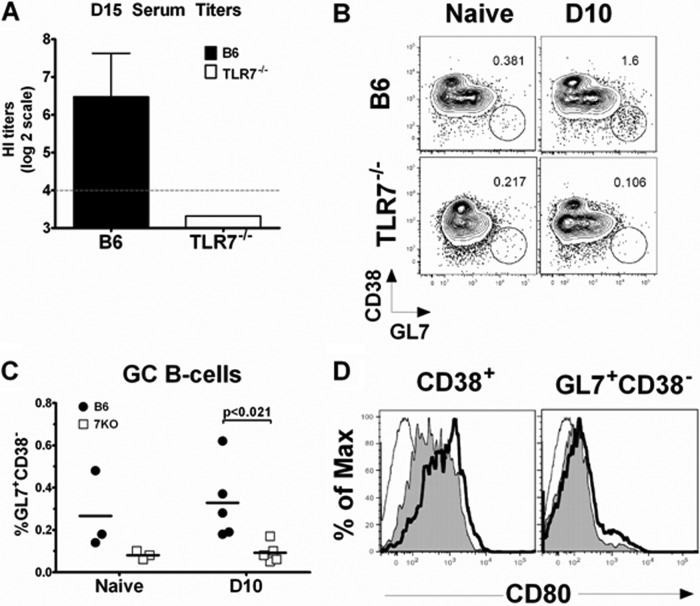
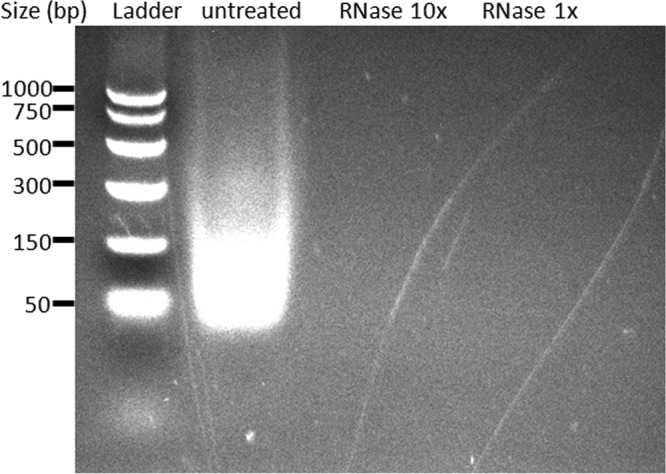
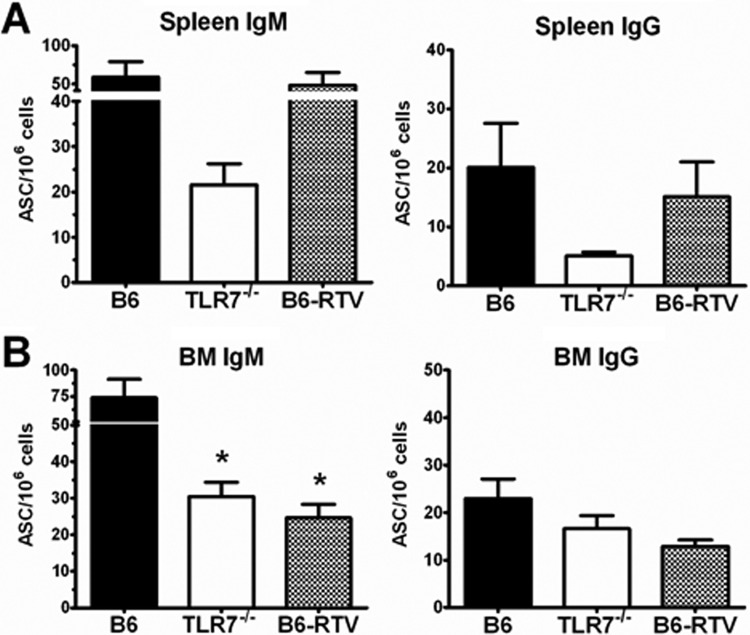
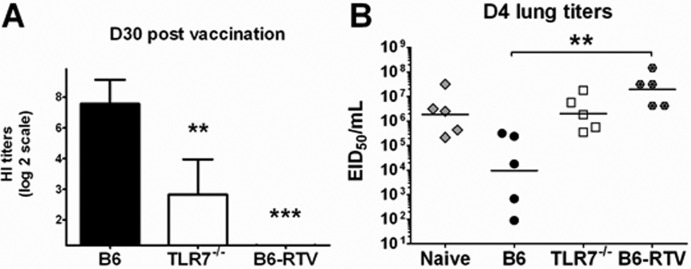
Recognition of pathogen-associated molecular patterns by pattern recognition receptors of the innate immune system is crucial for the initiation of innate and adaptive responses and for immunological memory. We investigated the role of TLR7 in the induction of adaptive immunity and long-term memory following influenza virus infection and vaccination in C57BL/6 mice. During infection with influenza A/PR8/34 virus, the absence of either TLR7 or MyD88 leads to reduced virus-specific antibodies in the serum and antibody-secreting cells in their secondary lymphoid organs, particularly in bone marrow. In spite of this, the absence of TLR7/MyD88 signaling did not impair the production of protective antibodies. Following immunization with the 2009 pandemic inactivated split vaccine, TLR7(-/-) mice had significantly lower levels of germinal center formation, antibody-secreting cells, and circulating influenza virus-specific antibodies than control animals. Consequently, TLR7(-/-) mice failed to develop protective immunological memory upon challenge. Furthermore, the immunogenicity of the split vaccine was likely due to TLR7 recognition of virion RNA, as its removal from the split vaccine significantly reduced the levels of influenza virus-specific antibodies and compromised the vaccine protective efficacy in mice. Taken together, our data demonstrate that TLR7 plays an important role in vaccine-induced humoral immune responses to influenza virus through the interaction with viral RNA present in the split vaccine.
Figures
Similar articles
-
Synthetic Toll-Like Receptor 4 (TLR4) and TLR7 Ligands Work Additively via MyD88 To Induce Protective Antiviral Immunity in Mice.J Virol. 2017 Sep 12;91(19):e01050-17. doi: 10.1128/JVI.01050-17. Print 2017 Oct 1. J Virol. 2017. PMID: 28724768 Free PMC article.
-
MyD88 signaling is indispensable for primary influenza A virus infection but dispensable for secondary infection.J Virol. 2010 Dec;84(24):12713-22. doi: 10.1128/JVI.01675-10. Epub 2010 Oct 13. J Virol. 2010. PMID: 20943980 Free PMC article.
-
Synthetic Toll-like receptor 4 (TLR4) and TLR7 ligands as influenza virus vaccine adjuvants induce rapid, sustained, and broadly protective responses.J Virol. 2015 Mar;89(6):3221-35. doi: 10.1128/JVI.03337-14. Epub 2015 Jan 7. J Virol. 2015. PMID: 25568203 Free PMC article.
-
Implications of broadly neutralizing antibodies in the development of a universal influenza vaccine.Curr Opin Virol. 2016 Apr;17:110-115. doi: 10.1016/j.coviro.2016.03.002. Epub 2016 Mar 28. Curr Opin Virol. 2016. PMID: 27031684 Free PMC article. Review.
-
Antibody Responsiveness to Influenza: What Drives It?Viruses. 2021 Jul 19;13(7):1400. doi: 10.3390/v13071400. Viruses. 2021. PMID: 34372607 Free PMC article. Review.
Cited by
-
Cytokine production in whole-blood cultures following immunization with an influenza vaccine.Hum Vaccin Immunother. 2018;14(12):2990-2998. doi: 10.1080/21645515.2018.1498435. Epub 2018 Aug 28. Hum Vaccin Immunother. 2018. PMID: 30036123 Free PMC article.
-
Biological sex affects vaccine efficacy and protection against influenza in mice.Proc Natl Acad Sci U S A. 2018 Dec 4;115(49):12477-12482. doi: 10.1073/pnas.1805268115. Epub 2018 Nov 19. Proc Natl Acad Sci U S A. 2018. PMID: 30455317 Free PMC article.
-
The battle between influenza and the innate immune response in the human respiratory tract.Infect Chemother. 2013 Mar;45(1):11-21. doi: 10.3947/ic.2013.45.1.11. Epub 2013 Mar 29. Infect Chemother. 2013. PMID: 24265946 Free PMC article. Review.
-
Induction and Subversion of Human Protective Immunity: Contrasting Influenza and Respiratory Syncytial Virus.Front Immunol. 2018 Mar 2;9:323. doi: 10.3389/fimmu.2018.00323. eCollection 2018. Front Immunol. 2018. PMID: 29552008 Free PMC article. Review.
-
B Cell Activation and Response Regulation During Viral Infections.Viral Immunol. 2020 May;33(4):294-306. doi: 10.1089/vim.2019.0207. Epub 2020 Apr 23. Viral Immunol. 2020. PMID: 32326852 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
