

Effects of drought on gene expression in maize reproductive and leaf meristem tissue revealed by RNA-Seq
- PMID: 22837360
- PMCID: PMC3461560
- DOI: 10.1104/pp.112.200444
Effects of drought on gene expression in maize reproductive and leaf meristem tissue revealed by RNA-Seq
Abstract
Drought stress affects cereals especially during the reproductive stage. The maize (Zea mays) drought transcriptome was studied using RNA-Seq analysis to compare drought-treated and well-watered fertilized ovary and basal leaf meristem tissue. More drought-responsive genes responded in the ovary compared with the leaf meristem. Gene Ontology enrichment analysis revealed a massive decrease in transcript abundance of cell division and cell cycle genes in the drought-stressed ovary only. Among Gene Ontology categories related to carbohydrate metabolism, changes in starch and Suc metabolism-related genes occurred in the ovary, consistent with a decrease in starch levels, and in Suc transporter function, with no comparable changes occurring in the leaf meristem. Abscisic acid (ABA)-related processes responded positively, but only in the ovaries. Related responses suggested the operation of low glucose sensing in drought-stressed ovaries. The data are discussed in the context of the susceptibility of maize kernel to drought stress leading to embryo abortion and the relative robustness of dividing vegetative tissue taken at the same time from the same plant subjected to the same conditions. Our working hypothesis involves signaling events associated with increased ABA levels, decreased glucose levels, disruption of ABA/sugar signaling, activation of programmed cell death/senescence through repression of a phospholipase C-mediated signaling pathway, and arrest of the cell cycle in the stressed ovary at 1 d after pollination. Increased invertase levels in the stressed leaf meristem, on the other hand, resulted in that tissue maintaining hexose levels at an "unstressed" level, and at lower ABA levels, which was correlated with successful resistance to drought stress.
Figures
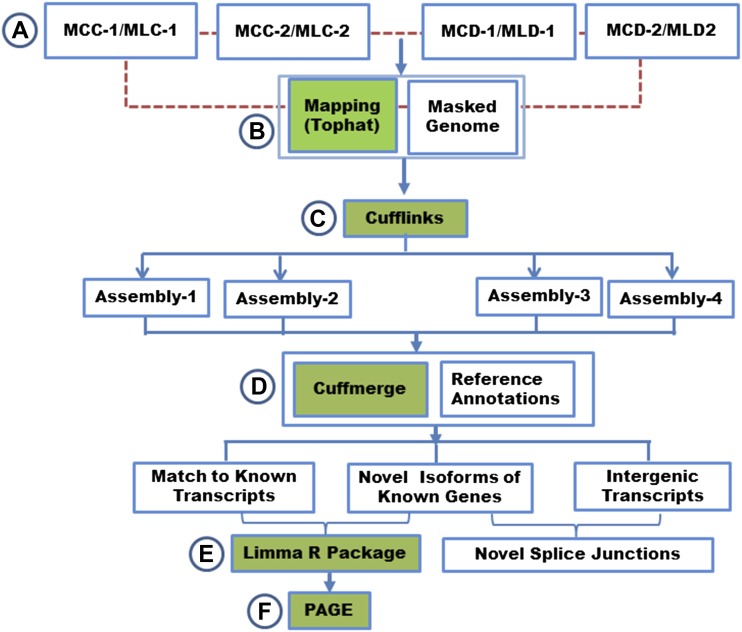
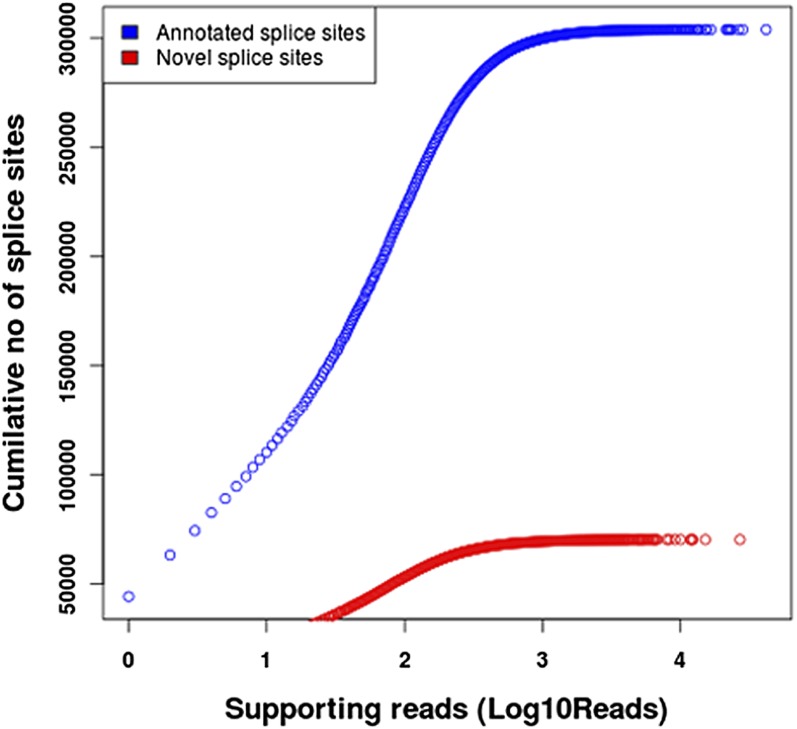


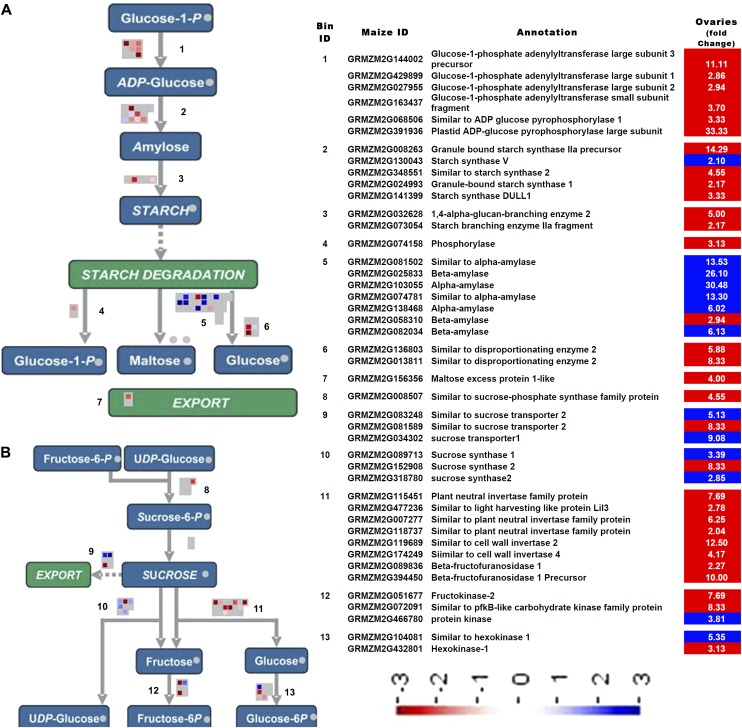
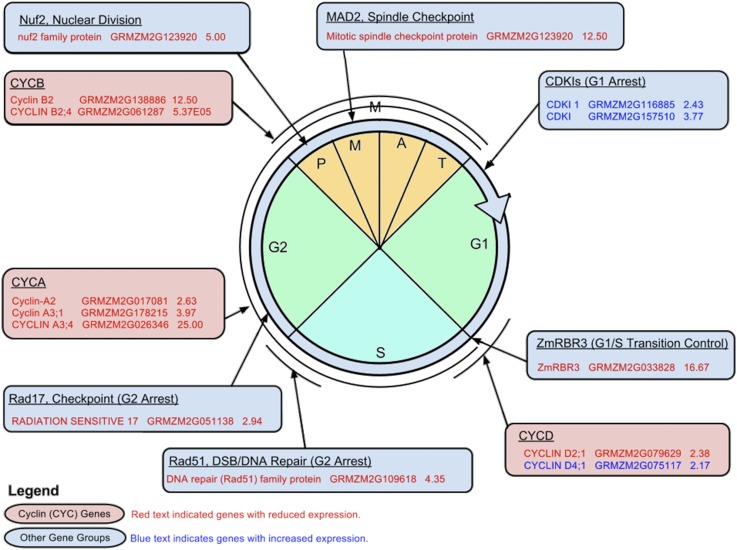
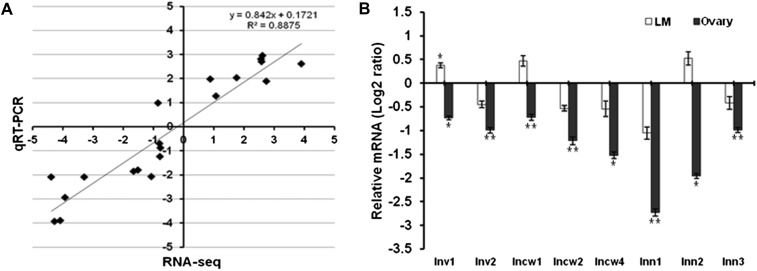
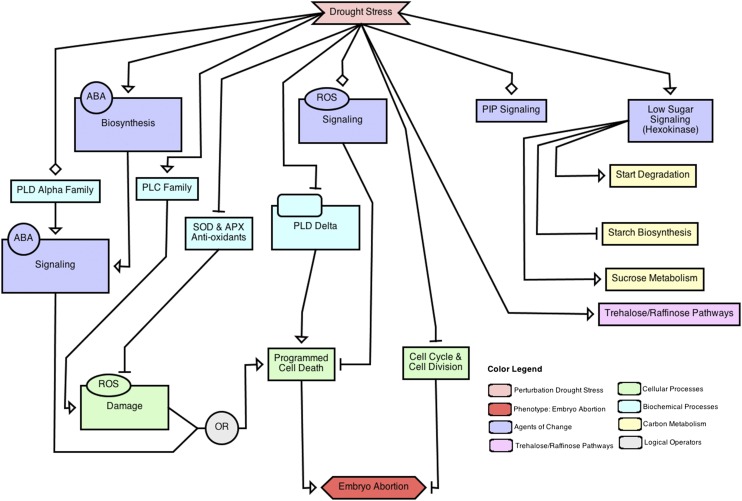
Similar articles
-
Key Maize Drought-Responsive Genes and Pathways Revealed by Comparative Transcriptome and Physiological Analyses of Contrasting Inbred Lines.Int J Mol Sci. 2019 Mar 13;20(6):1268. doi: 10.3390/ijms20061268. Int J Mol Sci. 2019. PMID: 30871211 Free PMC article.
-
Overexpression of the phosphatidylinositol synthase gene (ZmPIS) conferring drought stress tolerance by altering membrane lipid composition and increasing ABA synthesis in maize.Plant Cell Environ. 2013 May;36(5):1037-55. doi: 10.1111/pce.12040. Epub 2012 Dec 17. Plant Cell Environ. 2013. PMID: 23152961
-
Genetic association mapping identifies single nucleotide polymorphisms in genes that affect abscisic acid levels in maize floral tissues during drought.J Exp Bot. 2011 Jan;62(2):701-16. doi: 10.1093/jxb/erq308. Epub 2010 Nov 17. J Exp Bot. 2011. PMID: 21084430 Free PMC article.
-
Maize leaf yellowing gene ZmCAAX modulates growth and drought resistance by regulating abscisic acid contents through interaction with the ABA biosynthetic enzyme ZmNCED3.Plant Biotechnol J. 2025 Aug;23(8):3431-3450. doi: 10.1111/pbi.70147. Epub 2025 Jun 3. Plant Biotechnol J. 2025. PMID: 40458027 Free PMC article.
-
Transcriptional regulatory networks in response to drought stress and rewatering in maize (Zea mays L.).Mol Genet Genomics. 2021 Nov;296(6):1203-1219. doi: 10.1007/s00438-021-01820-y. Epub 2021 Oct 3. Mol Genet Genomics. 2021. PMID: 34601650 Review.
Cited by
-
New Insights on Drought Stress Response by Global Investigation of Gene Expression Changes in Sheepgrass (Leymus chinensis).Front Plant Sci. 2016 Jun 30;7:954. doi: 10.3389/fpls.2016.00954. eCollection 2016. Front Plant Sci. 2016. PMID: 27446180 Free PMC article.
-
Comparative transcriptomic analysis of contrasting hybrid cultivars reveal key drought-responsive genes and metabolic pathways regulating drought stress tolerance in maize at various stages.PLoS One. 2020 Oct 15;15(10):e0240468. doi: 10.1371/journal.pone.0240468. eCollection 2020. PLoS One. 2020. PMID: 33057352 Free PMC article.
-
The differential transcription network between embryo and endosperm in the early developing maize seed.Plant Physiol. 2013 May;162(1):440-55. doi: 10.1104/pp.113.214874. Epub 2013 Mar 11. Plant Physiol. 2013. PMID: 23478895 Free PMC article.
-
Genome-Wide Profiling of Alternative Splicing and Gene Fusion during Rice Black-Streaked Dwarf Virus Stress in Maize (Zea mays L.).Genes (Basel). 2022 Mar 2;13(3):456. doi: 10.3390/genes13030456. Genes (Basel). 2022. PMID: 35328010 Free PMC article.
-
Effective Use of Water in Crop Plants in Dryland Agriculture: Implications of Reactive Oxygen Species and Antioxidative System.Front Plant Sci. 2022 Jan 10;12:778270. doi: 10.3389/fpls.2021.778270. eCollection 2021. Front Plant Sci. 2022. PMID: 35082809 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
