

Origin of metazoan cadherin diversity and the antiquity of the classical cadherin/β-catenin complex
- PMID: 22837400
- PMCID: PMC3420208
- DOI: 10.1073/pnas.1120685109
Origin of metazoan cadherin diversity and the antiquity of the classical cadherin/β-catenin complex
Abstract
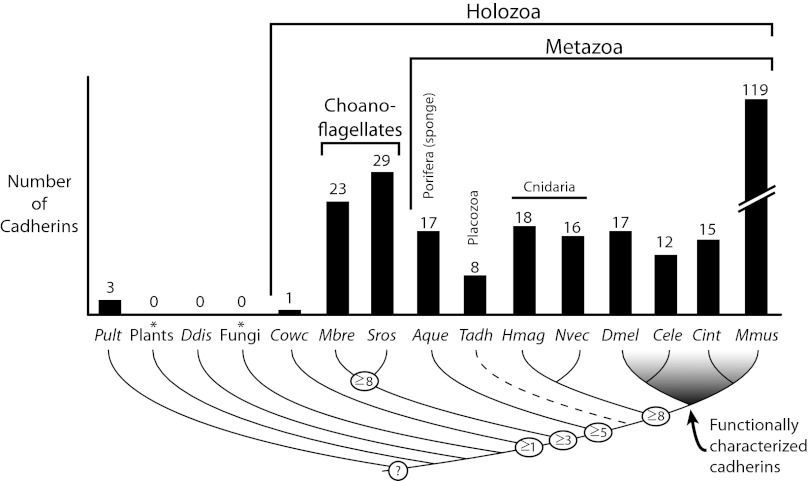
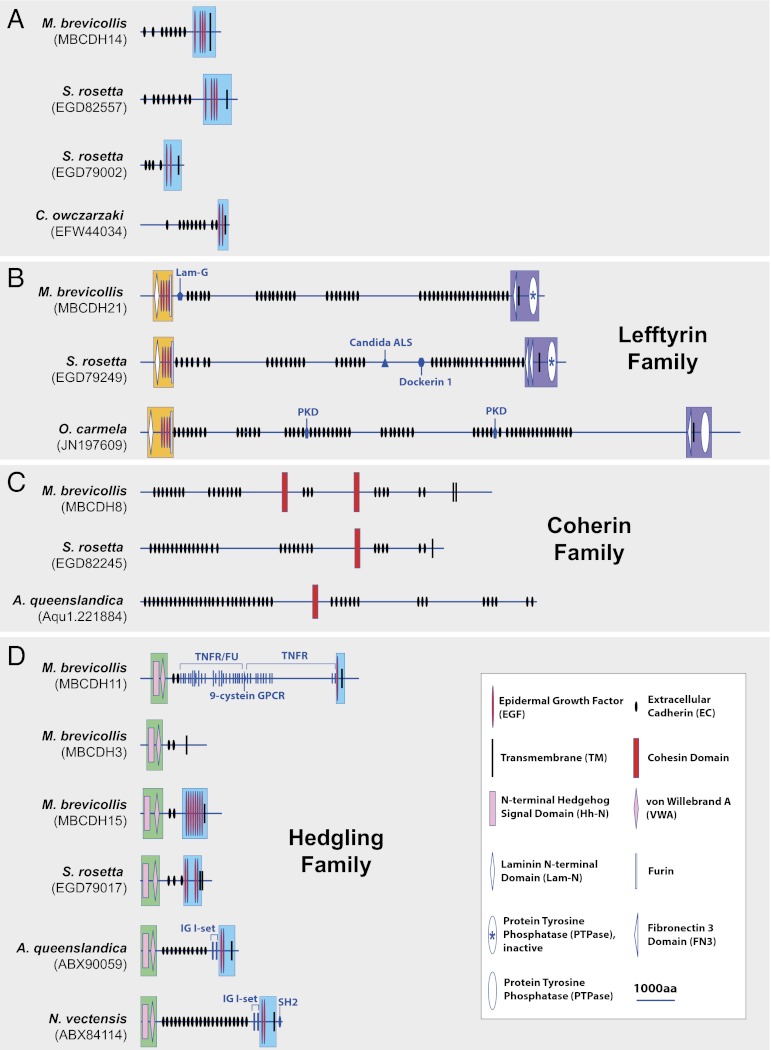
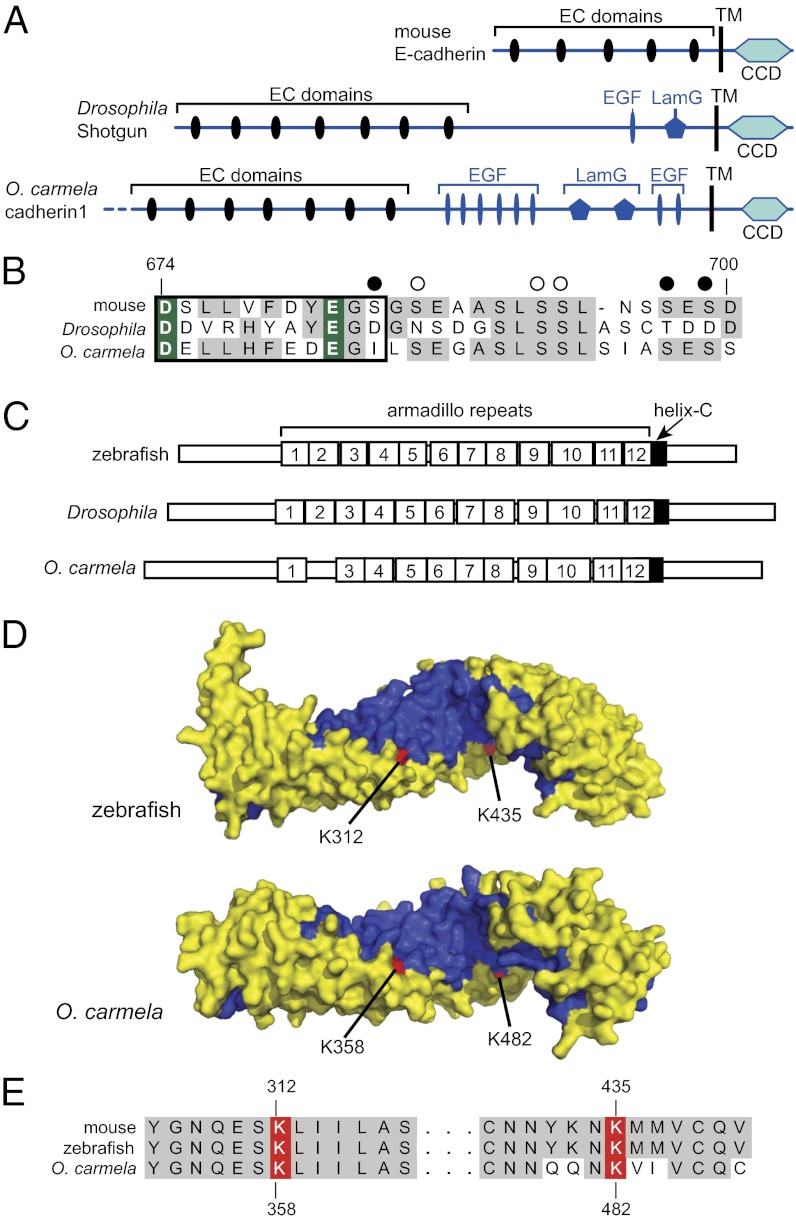
The evolution of cadherins, which are essential for metazoan multicellularity and restricted to metazoans and their closest relatives, has special relevance for understanding metazoan origins. To reconstruct the ancestry and evolution of cadherin gene families, we analyzed the genomes of the choanoflagellate Salpingoeca rosetta, the unicellular outgroup of choanoflagellates and metazoans Capsaspora owczarzaki, and a draft genome assembly from the homoscleromorph sponge Oscarella carmela. Our finding of a cadherin gene in C. owczarzaki reveals that cadherins predate the divergence of the C. owczarzaki, choanoflagellate, and metazoan lineages. Data from these analyses also suggest that the last common ancestor of metazoans and choanoflagellates contained representatives of at least three cadherin families, lefftyrin, coherin, and hedgling. Additionally, we find that an O. carmela classical cadherin has predicted structural features that, in bilaterian classical cadherins, facilitate binding to the cytoplasmic protein β-catenin and, thereby, promote cadherin-mediated cell adhesion. In contrast with premetazoan cadherin families (i.e., those conserved between choanoflagellates and metazoans), the later appearance of classical cadherins coincides with metazoan origins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
The Capsaspora genome reveals a complex unicellular prehistory of animals.Nat Commun. 2013;4:2325. doi: 10.1038/ncomms3325. Nat Commun. 2013. PMID: 23942320 Free PMC article.
-
The predicted secretomes of Monosiga brevicollis and Capsaspora owczarzaki, close unicellular relatives of metazoans, reveal new insights into the evolution of the metazoan extracellular matrix.Matrix Biol. 2014 Jul;37:60-8. doi: 10.1016/j.matbio.2014.02.002. Epub 2014 Feb 21. Matrix Biol. 2014. PMID: 24561726
-
Premetazoan genome evolution and the regulation of cell differentiation in the choanoflagellate Salpingoeca rosetta.Genome Biol. 2013 Feb 18;14(2):R15. doi: 10.1186/gb-2013-14-2-r15. Genome Biol. 2013. PMID: 23419129 Free PMC article.
-
Choanoflagellate models - Monosiga brevicollis and Salpingoeca rosetta.Curr Opin Genet Dev. 2016 Aug;39:42-47. doi: 10.1016/j.gde.2016.05.016. Epub 2016 Jun 17. Curr Opin Genet Dev. 2016. PMID: 27318693 Review.
-
Evolution and diversity of cadherins and catenins.Exp Cell Res. 2017 Sep 1;358(1):3-9. doi: 10.1016/j.yexcr.2017.03.001. Epub 2017 Mar 6. Exp Cell Res. 2017. PMID: 28268172 Review.
Cited by
-
Tracing the Evolutionary Origin of Desmosomes.Curr Biol. 2020 May 18;30(10):R535-R543. doi: 10.1016/j.cub.2020.03.047. Curr Biol. 2020. PMID: 32428495 Free PMC article. Review.
-
Evolutionary insights into premetazoan functions of the neuronal protein homer.Mol Biol Evol. 2014 Sep;31(9):2342-55. doi: 10.1093/molbev/msu178. Epub 2014 Jun 3. Mol Biol Evol. 2014. PMID: 24899667 Free PMC article.
-
The Desmosome-Keratin Scaffold Integrates ErbB Family and Mechanical Signaling to Polarize Epidermal Structure and Function.Front Cell Dev Biol. 2022 May 24;10:903696. doi: 10.3389/fcell.2022.903696. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35686051 Free PMC article. Review.
-
SAX-7/L1CAM acts with the adherens junction proteins MAGI-1, HMR-1/Cadherin, and AFD-1/Afadin to promote glial-mediated dendrite extension.bioRxiv [Preprint]. 2024 Jan 11:2024.01.11.575259. doi: 10.1101/2024.01.11.575259. bioRxiv. 2024. PMID: 38260503 Free PMC article. Preprint.
-
Co-expression of synaptic genes in the sponge Amphimedon queenslandica uncovers ancient neural submodules.Sci Rep. 2019 Oct 31;9(1):15781. doi: 10.1038/s41598-019-51282-x. Sci Rep. 2019. PMID: 31673079 Free PMC article.
References
-
- Abedin M, King N. The premetazoan ancestry of cadherins. Science. 2008;319:946–948. - PubMed
-
- Hulpiau P, van Roy F. New insights into the evolution of metazoan cadherins. Mol Biol Evol. 2011;28:647–657. - PubMed
-
- Hynes RO, Zhao Q. The evolution of cell adhesion. J Cell Biol. 2000;150:F89–F96. - PubMed
-
- Rokas A. The origins of multicellularity and the early history of the genetic toolkit for animal development. Annu Rev Genet. 2008;42:235–251. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
