

CD4+ T cell-dependent IFN-γ production by CD8+ effector T cells in Mycobacterium tuberculosis infection
- PMID: 22837486
- PMCID: PMC3424308
- DOI: 10.4049/jimmunol.1200994
CD4+ T cell-dependent IFN-γ production by CD8+ effector T cells in Mycobacterium tuberculosis infection
Abstract
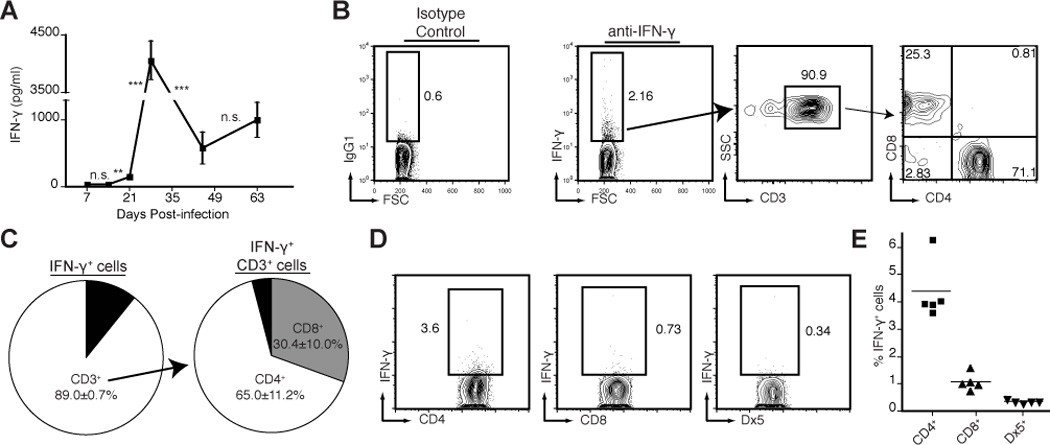
Both CD4+ and CD8+ T cells contribute to immunity to tuberculosis, and both can produce the essential effector cytokine IFN-γ. However, the precise role and relative contribution of each cell type to in vivo IFN-γ production are incompletely understood. To identify and quantitate the cells that produce IFN-γ at the site of Mycobacterium tuberculosis infection in mice, we used direct intracellular cytokine staining ex vivo without restimulation. We found that CD4+ and CD8+ cells were predominantly responsible for production of this cytokine in vivo, and we observed a remarkable linear correlation between the fraction of CD4+ cells and the fraction of CD8+ cells producing IFN-γ in the lungs. In the absence of CD4+ cells, a reduced fraction of CD8+ cells was actively producing IFN-γ in vivo, suggesting that CD4+ effector cells are continually required for optimal IFN-γ production by CD8+ effector cells. Accordingly, when infected mice were treated i.v. with an MHC-II-restricted M. tuberculosis epitope peptide to stimulate CD4+ cells in vivo, we observed rapid activation of both CD4+ and CD8+ cells in the lungs. Indirect activation of CD8+ cells was dependent on the presence of CD4+ cells but independent of IFN-g responsiveness of the CD8+ cells. These data provide evidence that CD4+ cell deficiency impairs IFN-γ production by CD8+ effector cells and that ongoing cross-talk between distinct effector T cell types in the lungs may contribute to a protective immune response against M. tuberculosis. Conversely, defects in these interactions may contribute to susceptibility to tuberculosis and other infections.
Conflict of interest statement
The authors have no conflicting financial interests.
Figures




References
-
- Filipe-Santos O, Bustamante J, Chapgier A, Vogt G, de Beaucoudrey L, Feinberg J, Jouanguy E, Boisson-Dupuis S, Fieschi C, Picard C, Casanova JL. Inborn errors of IL-12/23- and IFN-gamma-mediated immunity: molecular, cellular, and clinical features. Semin Immunol. 2006;18:347–361. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
