

Silencing of Apoptosis-Inducing factor and poly(ADP-ribose) glycohydrolase reveals novel roles in breast cancer cell death after chemotherapy
- PMID: 22839996
- PMCID: PMC3494550
- DOI: 10.1186/1476-4598-11-48
Silencing of Apoptosis-Inducing factor and poly(ADP-ribose) glycohydrolase reveals novel roles in breast cancer cell death after chemotherapy
Abstract
Background: Cell death induced by poly(ADP-ribose) (PAR) and mediated by apoptosis-inducing factor (AIF) is well-characterized in models of ischemic tissue injury, but their roles in cancer cell death after chemotherapy are less understood.
Methods: Here we investigated the roles of PAR and AIF by RNA interference (RNAi) in MDA-MB-231 and MCF-7 breast adenocarcinoma cells after chemotherapy. Differences in effects were statistically tested by analysis-of-variance and unpaired student's t-test.
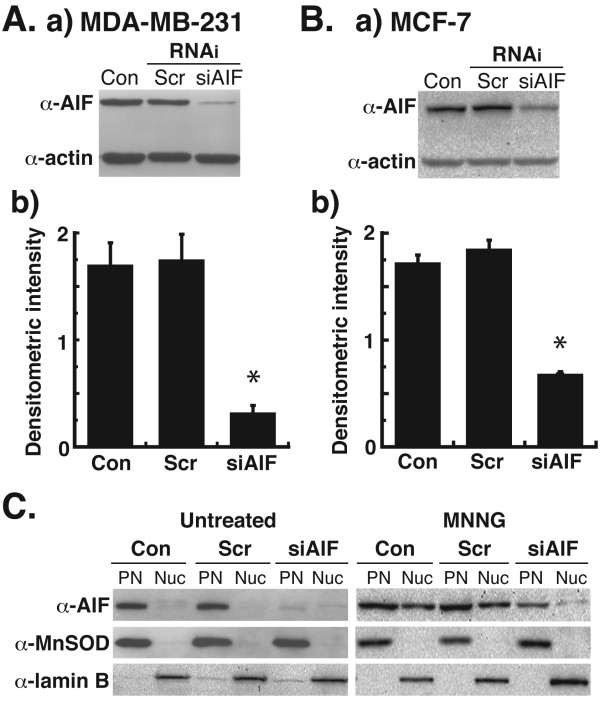
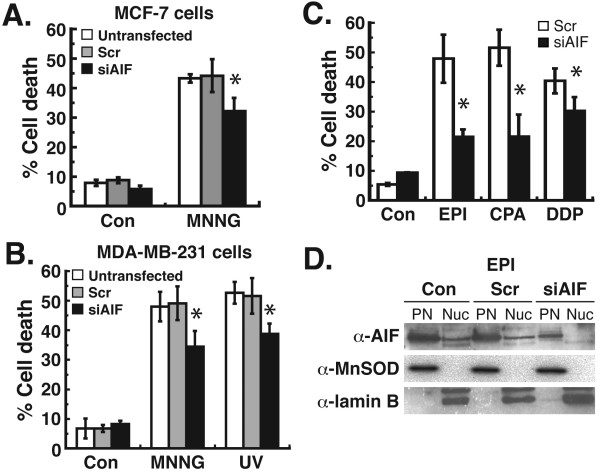
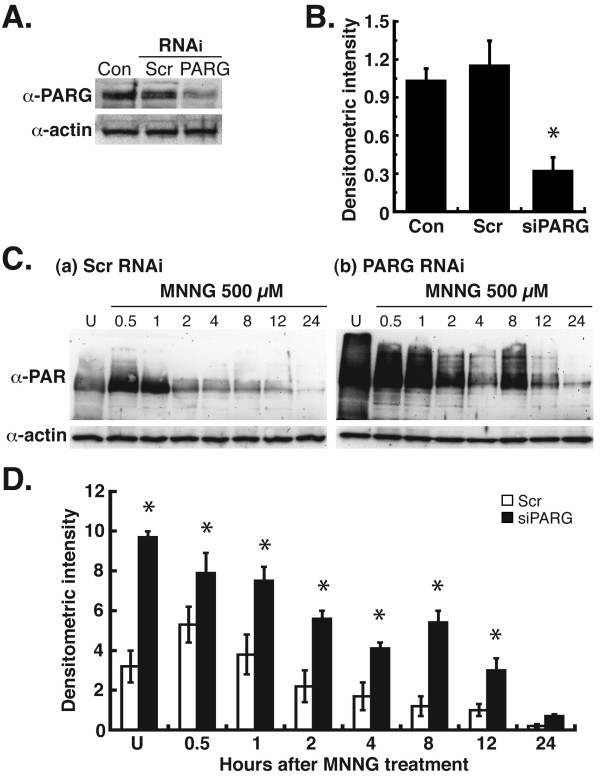
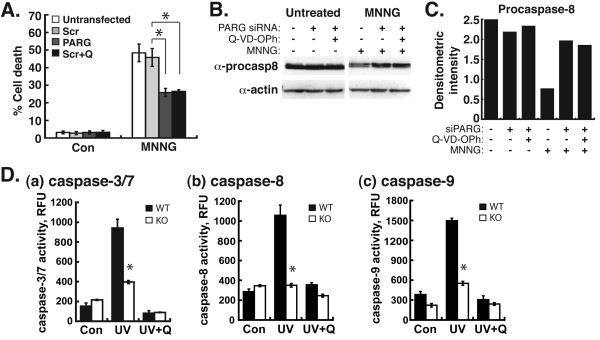
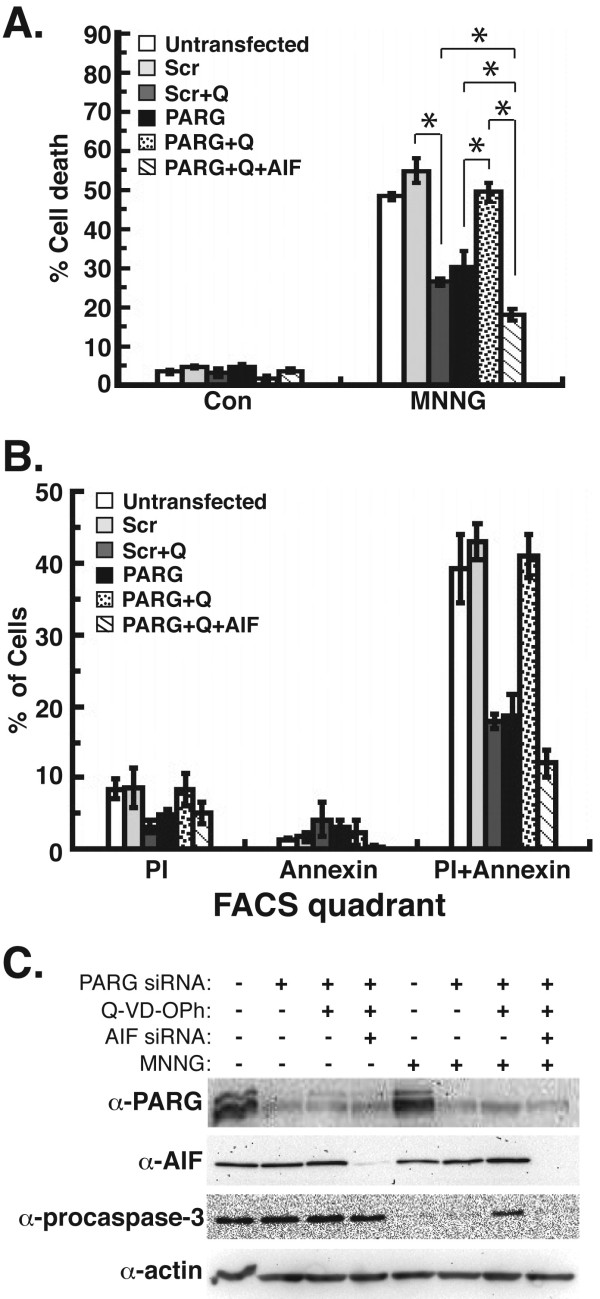
Results: Silencing of AIF by RNAi led to decreased MDA-MB-231 and MCF-7 breast cancer cell death after chemotherapy, which demonstrates a critical role for AIF. RNAi silencing of PAR glycohydrolase (PARG), the primary enzyme that catalyzes the hydrolysis of PAR, led to increased PAR levels but decreased cell death. Further investigation into the possible role of PAR in apoptosis revealed decreased caspase-3/7/8/9 activity in PARG-null cells. Interestingly, the pharmacologic inhibition of caspase activity in PARG-silenced breast cancer cells led to increased cell death after chemotherapy, which indicates that an alternative cell death pathway is activated due to elevated PAR levels and caspase inhibition. AIF silencing in these cells led to profound protection from chemotherapy, which demonstrates that the increased cell death after PARG silencing and caspase inhibition was mediated by AIF.
Conclusions: The results show a role for AIF in breast cancer cell death after chemotherapy, the ability of PAR to regulate caspase activity, and the ability of AIF to substitute as a primary mediator of breast cancer cell death in the absence of caspases. Thus, the induction of cell death by PAR/AIF may represent a novel strategy to optimize the eradication of breast tumors by activating an alternative cell death pathway.
Figures
References
-
- Pospisilik JA, Knauf C, Joza N, Benit P, Orthofer M, Cani PD, Ebersberger I, Nakashima T, Sarao R, Neely G. et al.Targeted deletion of AIF decreases mitochondrial oxidative phosphorylation and protects from obesity and diabetes. Cell. 2007;131:476–491. doi: 10.1016/j.cell.2007.08.047. - DOI - PubMed
-
- Artus C, Boujrad H, Bouharrour A, Brunelle MN, Hoos S, Yuste VJ, Lenormand P, Rousselle JC, Namane A, England P. et al.AIF promotes chromatinolysis and caspase-independent programmed necrosis by interacting with histone H2AX. EMBO J. 2010;29:1585–1599. doi: 10.1038/emboj.2010.43. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
