

Quantitative live cell imaging reveals a gradual shift between DNA repair mechanisms and a maximal use of HR in mid S phase
- PMID: 22841003
- PMCID: PMC3494418
- DOI: 10.1016/j.molcel.2012.05.052
Quantitative live cell imaging reveals a gradual shift between DNA repair mechanisms and a maximal use of HR in mid S phase
Abstract
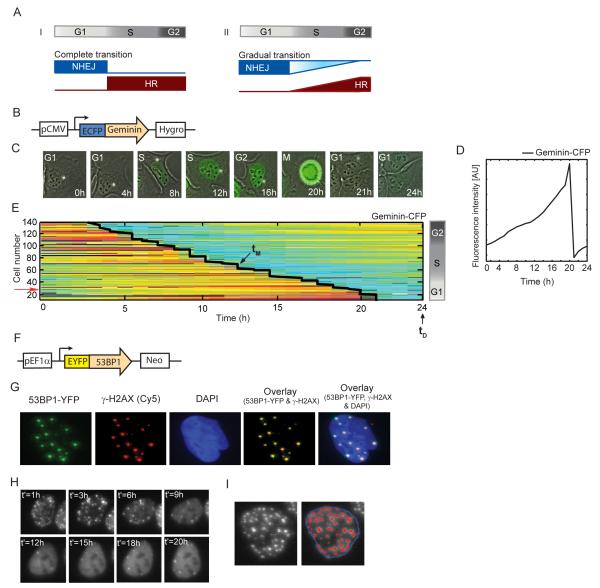
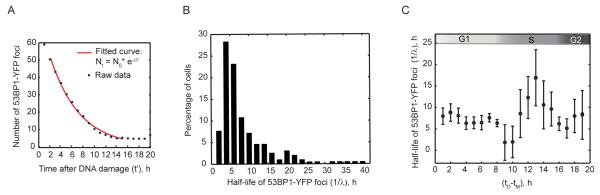
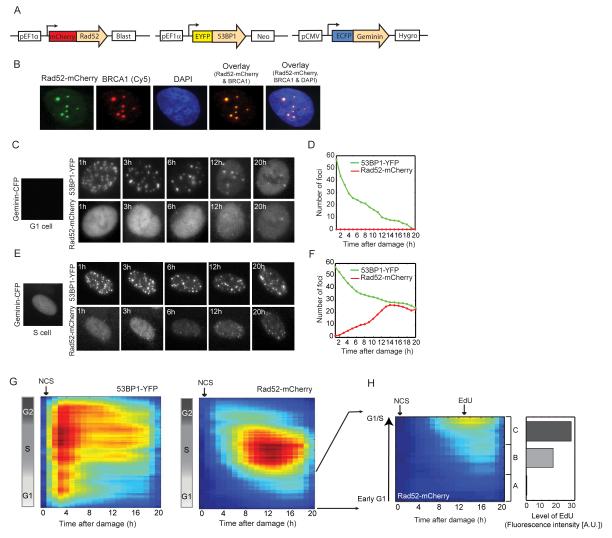
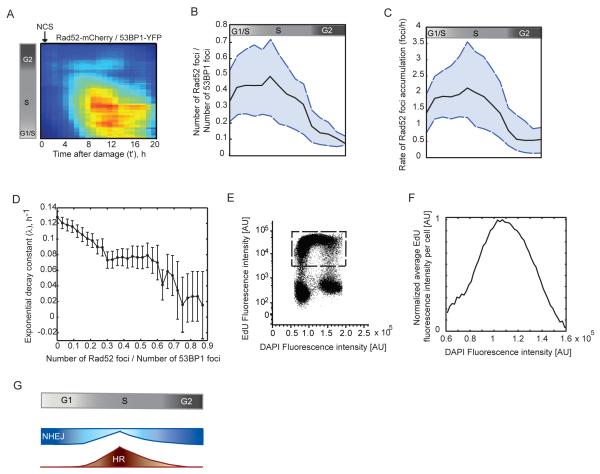
DNA double-strand breaks are repaired by two main pathways: nonhomologous end joining (NHEJ) and homologous recombination (HR). The choice between these pathways depends on cell-cycle phase; however the continuous effect of cell cycle on the balance between them is still unclear. We used live cell imaging and fluorescent reporters for 53BP1, Rad52, and cell cycle to quantify the relative contribution of NHEJ and HR at different points of the cell cycle in single cells. We found that NHEJ is the dominant repair pathway in G1 and G2 even when both repair pathways are functional. The shift from NHEJ to HR is gradual, with the highest proportion of breaks repaired by HR in mid S, where the amount of DNA replication is highest. Higher proportions of HR also strongly correlate with slower rates of repair. Our study shows that the choice of repair mechanism is continuously adjusted throughout the cell cycle and suggests that the extent of active replication, rather than the presence of a sister chromatid influences the balance between the two repair pathways in human cells.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
