

Mechanisms of neuronal computation in mammalian visual cortex
- PMID: 22841306
- PMCID: PMC3477598
- DOI: 10.1016/j.neuron.2012.06.011
Mechanisms of neuronal computation in mammalian visual cortex
Abstract
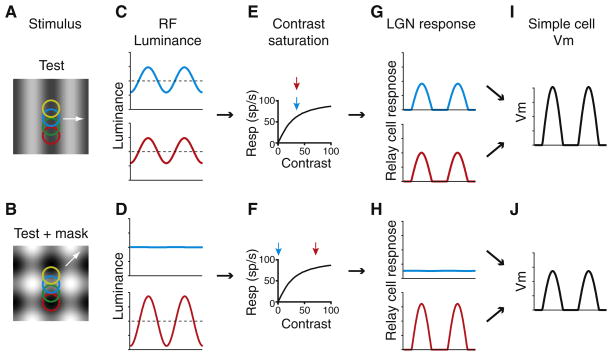
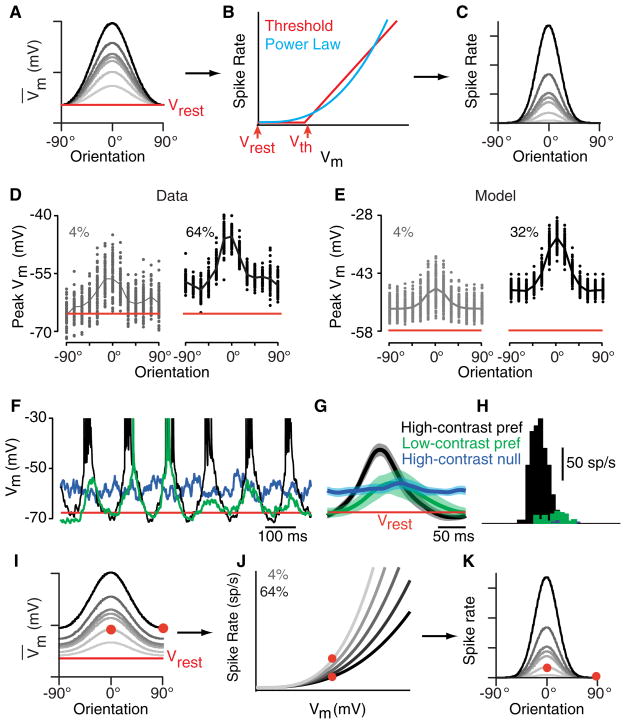
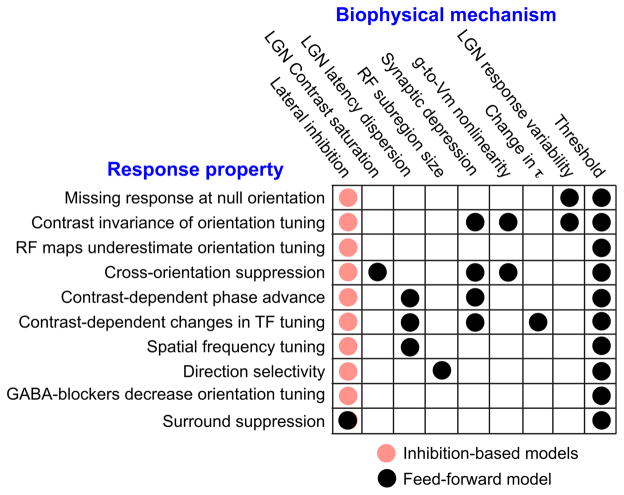
Orientation selectivity in the primary visual cortex (V1) is a receptive field property that is at once simple enough to make it amenable to experimental and theoretical approaches and yet complex enough to represent a significant transformation in the representation of the visual image. As a result, V1 has become an area of choice for studying cortical computation and its underlying mechanisms. Here we consider the receptive field properties of the simple cells in cat V1--the cells that receive direct input from thalamic relay cells--and explore how these properties, many of which are highly nonlinear, arise. We have found that many receptive field properties of V1 simple cells fall directly out of Hubel and Wiesel's feedforward model when the model incorporates realistic neuronal and synaptic mechanisms, including threshold, synaptic depression, response variability, and the membrane time constant.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures





References
-
- Albrecht DG. Visual cortex neurons in monkey and cat: effect of contrast on the spatial and temporal phase transfer functions. Vis Neurosci. 1995;12:1191–1210. - PubMed
-
- Alitto HJ, Usrey WM. Influence of contrast on orientation and temporal frequency tuning in ferret primary visual cortex. J Neurophysiol. 2004;91:2797–2808. - PubMed
-
- Alonso JM, Martinez LM. Functional connectivity between simple cells and complex cells in cat striate cortex. Nat Neurosci. 1998;1:395–403. - PubMed
-
- Anderson JS, Carandini M, Ferster D. Orientation tuning of input conductance, excitation, and inhibition in cat primary visual cortex. J Neurophysiol. 2000;84:909–926. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
