

Photochemical restoration of visual responses in blind mice
- PMID: 22841312
- PMCID: PMC3408583
- DOI: 10.1016/j.neuron.2012.05.022
Photochemical restoration of visual responses in blind mice
Abstract
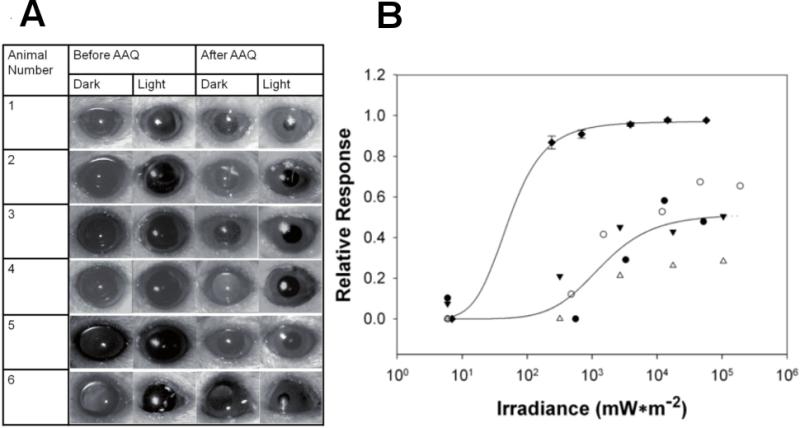
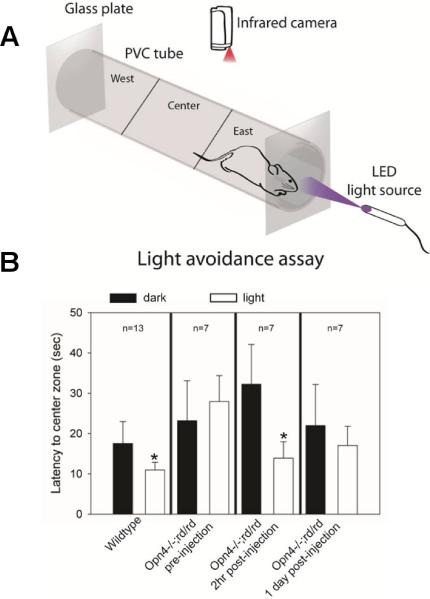
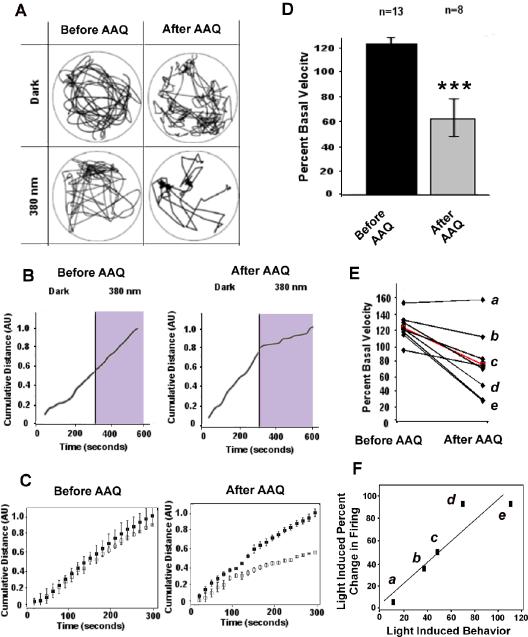
Retinitis pigmentosa (RP) and age-related macular degeneration (AMD) are degenerative blinding diseases caused by the death of rods and cones, leaving the remainder of the visual system intact but largely unable to respond to light. Here, we show that AAQ, a synthetic small molecule photoswitch, can restore light sensitivity to the retina and behavioral responses in vivo in mouse models of RP, without exogenous gene delivery. Brief application of AAQ bestows prolonged light sensitivity on multiple types of retinal neurons, resulting in synaptically amplified responses and center-surround antagonism in arrays of retinal ganglion cells (RGCs). Intraocular injection of AAQ restores the pupillary light reflex and locomotory light avoidance behavior in mice lacking retinal photoreceptors, indicating reconstitution of light signaling to brain circuits. AAQ and related photoswitch molecules present a potential drug strategy for restoring retinal function in degenerative blinding diseases.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
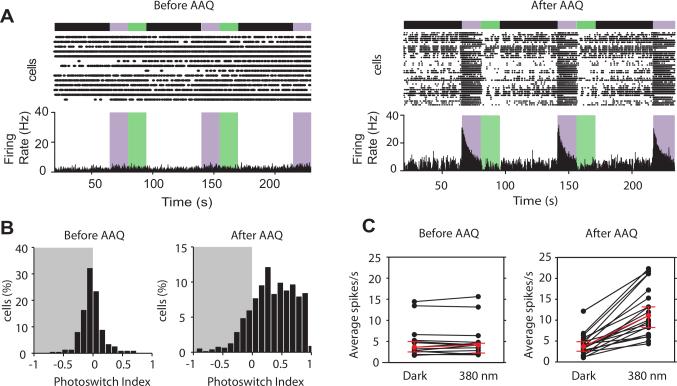
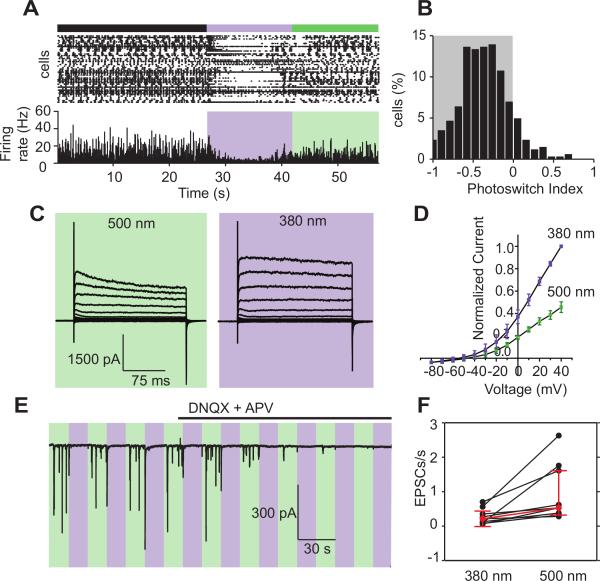
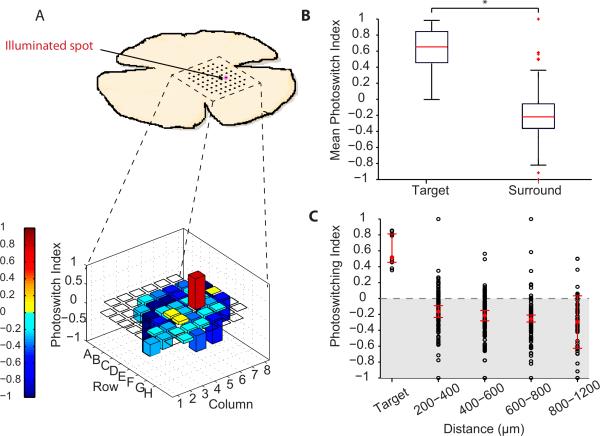
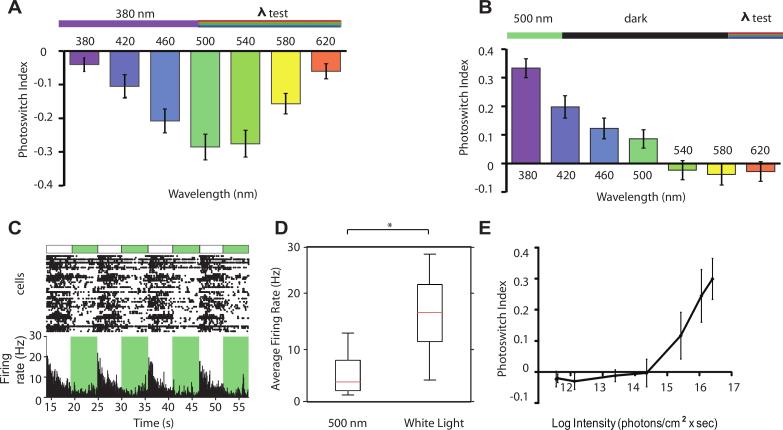
Comment in
-
The bionic retina: a small molecule with big potential for visual restoration.Neuron. 2012 Jul 26;75(2):185-7. doi: 10.1016/j.neuron.2012.06.010. Neuron. 2012. PMID: 22841303
References
-
- Benav H, Bartz-Schmidt KU, Besch D, Bruckmann A, Gekeler F, Greppmaier U, Harscher A, Kibbel S, Kusnyerik A, Peters T, et al. Restoration of useful vision up to letter recognition capabilities using subretinal microphotodiodes.. Engineering in Medicine and Biology Society (EMBC), 2010 Annual International Conference of the IEEE.2010. pp. 5919–5922. - PubMed
