

Copper influx transporter 1 is required for FGF, PDGF and EGF-induced MAPK signaling
- PMID: 22842628
- PMCID: PMC3464187
- DOI: 10.1016/j.bcp.2012.07.014
Copper influx transporter 1 is required for FGF, PDGF and EGF-induced MAPK signaling
Abstract
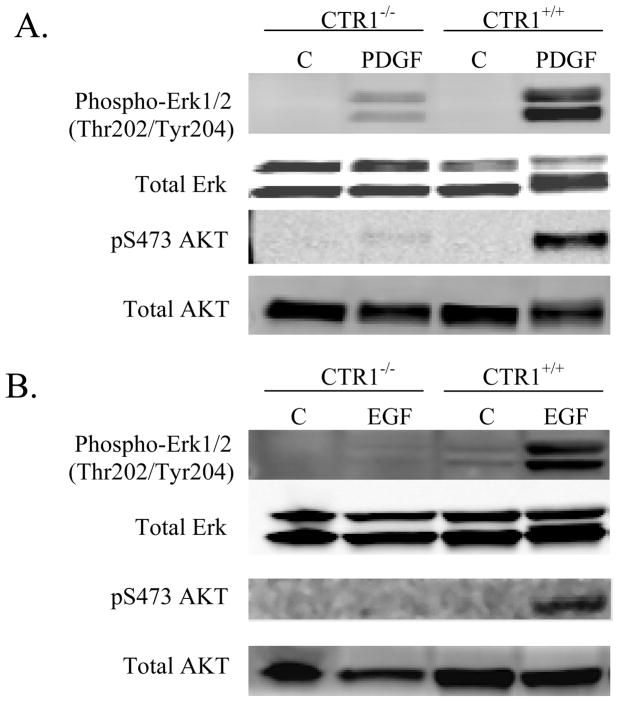
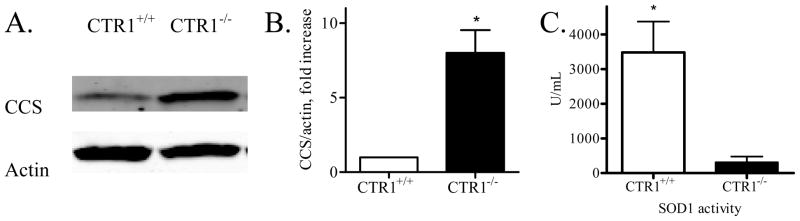
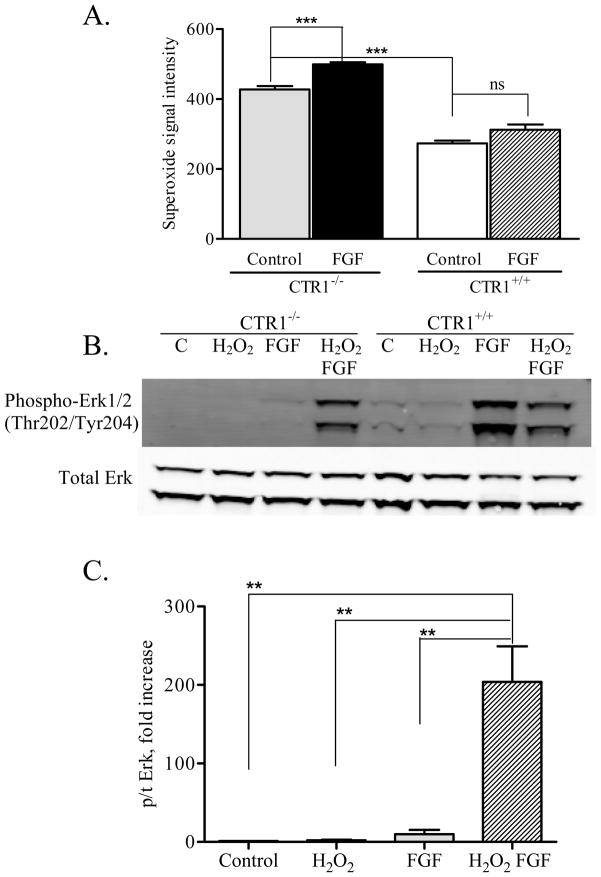
Copper transporter 1 (CTR1) is the major copper (Cu) influx transporter in mammalian cells. We report here that CTR1 is required for the activation of signaling to the MAPK pathway by the ligands of three major receptor tyrosine kinases (RTK) including FGF, PDGF and EGF. Induction of Erk1/2 phosphorylation was compared in isogenic wild type CTR1(+/+) and CTR1(-/-) cells. Whereas all three ligands increased pErk1/2 in the CTR1(+/+) cells, they failed to do this in CTR1(-/-) cells. While FGF did not enhance the phosphorylation of AKT in the CTR1(+/+) cells, both PDGF and EGF increased pAKT in the CTR1(+/+) but not CTR1(-/-) cells. The deficit in Erk1/2 phosphorylation in the CTR1(-/-) cells was rescued by adding Cu to the medium, and it was induced in CTR1(+/+) cells by treatment with a Cu chelator. Intracellular Cu availability was reduced in the CTR1(-/-) cells as reflected by increased expression of the Cu chaperone CCS. The failure of RTK-induced signaling to both Erk1/2 and AKT suggested the presence of a Cu-dependent step upstream of Ras. The Cu-dependent enzyme SOD1 is responsible for generating the hydrogen peroxide in response to RTK activation that serves to inhibit phosphatases that normally limit RTK signaling. SOD1 activity was reduced by a factor of 17-fold in the CTR1(-/-) cells, and addition of hydrogen peroxide restored signaling. We conclude that Cu acquired from CTR1 is required for signaling in pathways regulated by RTKs that play major roles in development and cancer.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures



References
-
- Rhee SG. Cell signaling. H2O2, a necessary evil for cell signaling. Science. 2006;312:1882–3. - PubMed
-
- Tonks NK. Redox redux: revisiting PTPs and the control of cell signaling. Cell. 2005;121:667–70. - PubMed
-
- Rhee SG, Kang SW, Jeong W, Chang TS, Yang KS, Woo HA. Intracellular messenger function of hydrogen peroxide and its regulation by peroxiredoxins. Curr Opin Cell Biol. 2005;17:183–9. - PubMed
-
- Ushio-Fukai M. Redox signaling in angiogenesis: role of NADPH oxidase. Cardiovascular research. 2006;71:226–35. - PubMed
-
- Rae TD, Schmidt PJ, Pufahl RA, Culotta VC, O’Halloran TV. Undetectable intracellular free copper: the requirement of a copper chaperone for superoxide dismutase. Science. 1999;284:805–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
