

Enhanced β-secretase processing alters APP axonal transport and leads to axonal defects
- PMID: 22843498
- PMCID: PMC3471392
- DOI: 10.1093/hmg/dds297
Enhanced β-secretase processing alters APP axonal transport and leads to axonal defects
Abstract
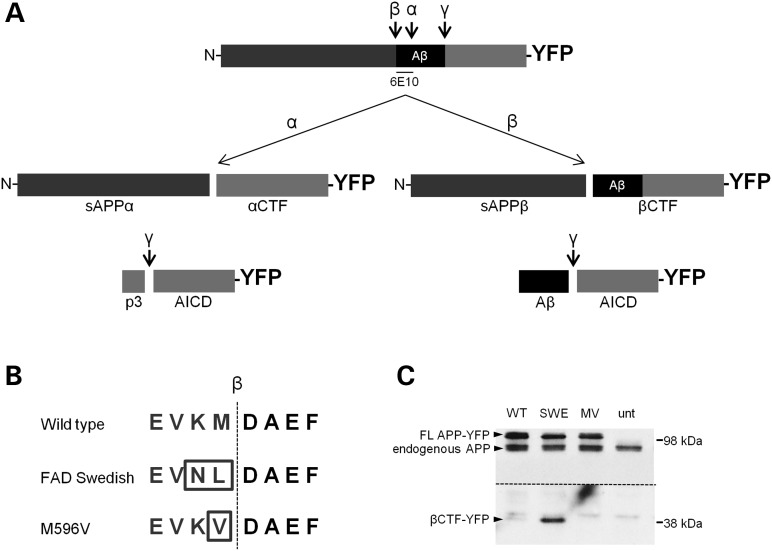
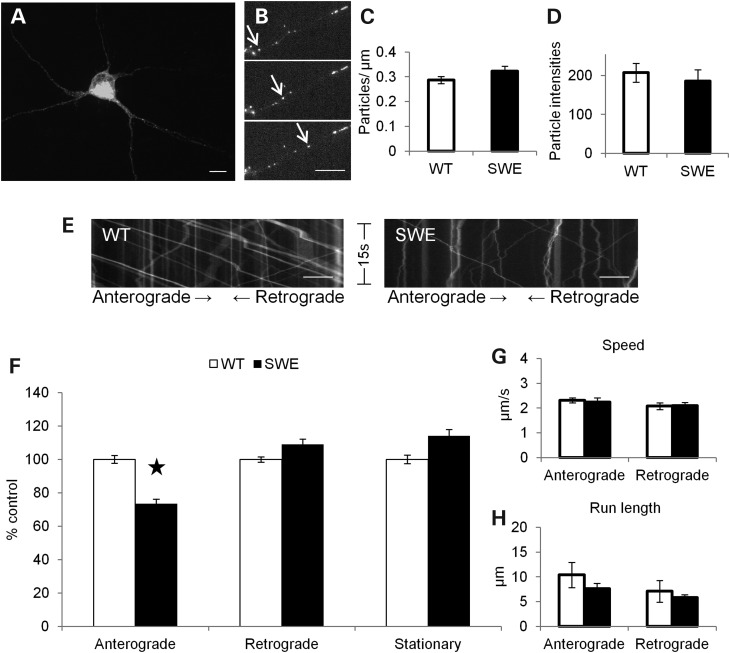
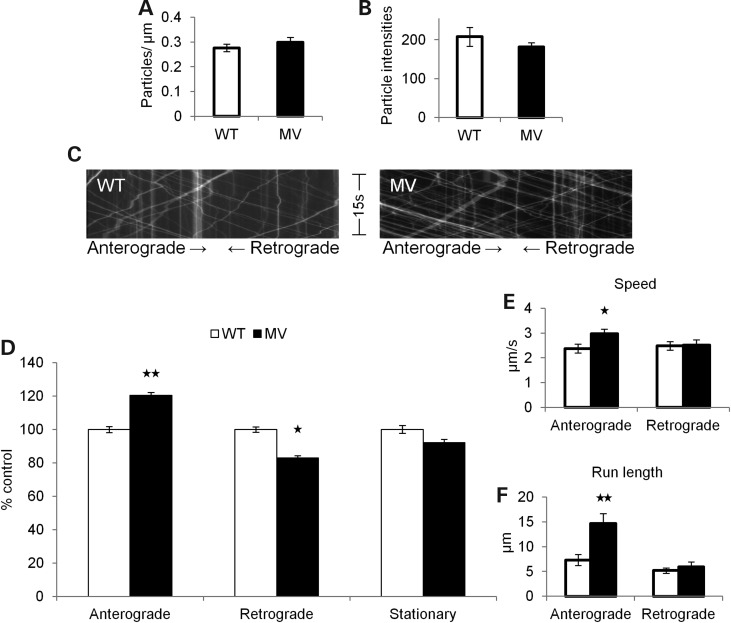
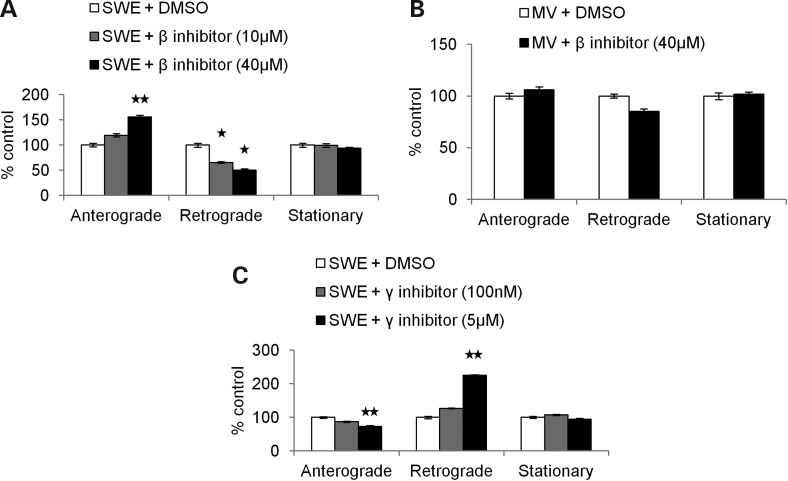
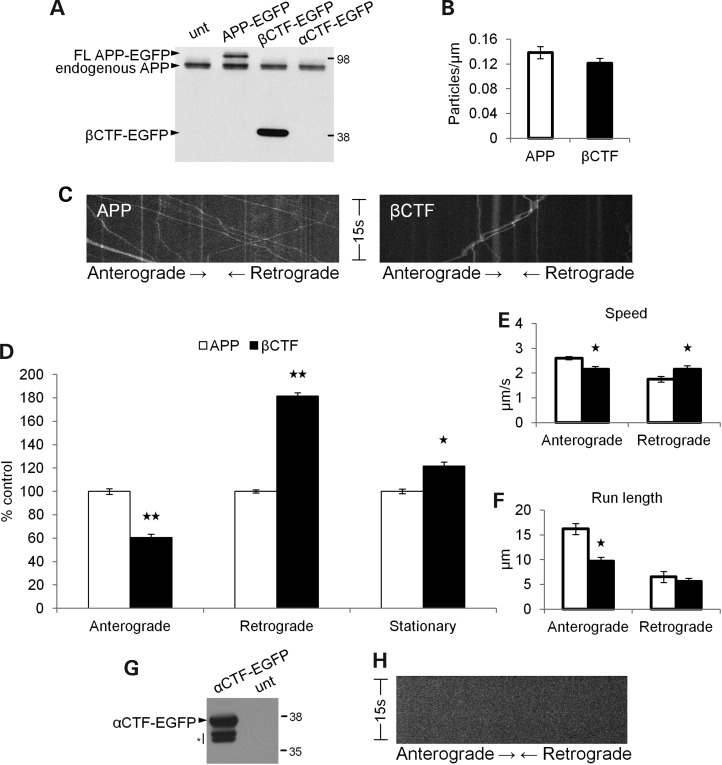
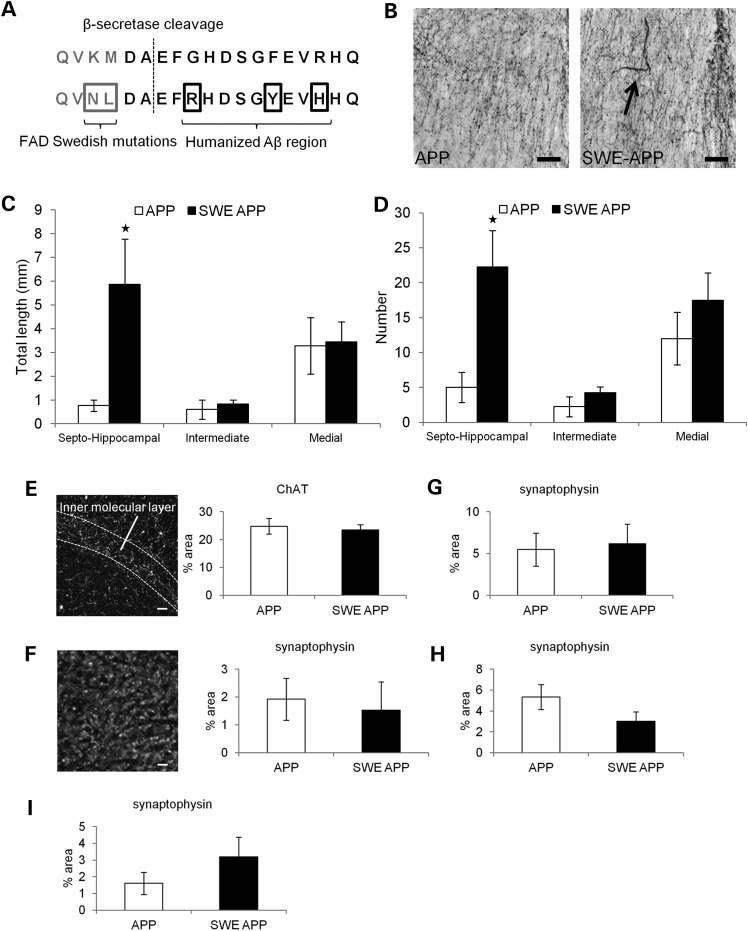
Alzheimer's disease (AD) is a neurodegenerative disease pathologically characterized by amyloid plaques and neurofibrillary tangles in the brain. Before these hallmark features appear, signs of axonal transport defects develop, though the initiating events are not clear. Enhanced amyloidogenic processing of amyloid precursor protein (APP) plays an integral role in AD pathogenesis, and previous work suggests that both the Aβ region and the C-terminal fragments (CTFs) of APP can cause transport defects. However, it remains unknown if APP processing affects the axonal transport of APP itself, and whether increased APP processing is sufficient to promote axonal dystrophy. We tested the hypothesis that β-secretase cleavage site mutations of APP alter APP axonal transport directly. We found that the enhanced β-secretase cleavage reduces the anterograde axonal transport of APP, while inhibited β-cleavage stimulates APP anterograde axonal transport. Transport behavior of APP after treatment with β- or γ-secretase inhibitors suggests that the amount of β-secretase cleaved CTFs (βCTFs) of APP underlies these transport differences. Consistent with these findings, βCTFs have reduced anterograde axonal transport compared with full-length, wild-type APP. Finally, a gene-targeted mouse with familial AD (FAD) Swedish mutations to APP, which enhance the β-cleavage of APP, develops axonal dystrophy in the absence of mutant protein overexpression, amyloid plaque deposition and synaptic degradation. These results suggest that the enhanced β-secretase processing of APP can directly impair the anterograde axonal transport of APP and are sufficient to lead to axonal defects in vivo.
Figures
Similar articles
-
Amyloid-β protein (Aβ) Glu11 is the major β-secretase site of β-site amyloid-β precursor protein-cleaving enzyme 1(BACE1), and shifting the cleavage site to Aβ Asp1 contributes to Alzheimer pathogenesis.Eur J Neurosci. 2013 Jun;37(12):1962-9. doi: 10.1111/ejn.12235. Eur J Neurosci. 2013. PMID: 23773065
-
Regulation of Synaptic Amyloid-β Generation through BACE1 Retrograde Transport in a Mouse Model of Alzheimer's Disease.J Neurosci. 2017 Mar 8;37(10):2639-2655. doi: 10.1523/JNEUROSCI.2851-16.2017. Epub 2017 Feb 3. J Neurosci. 2017. PMID: 28159908 Free PMC article.
-
Amyloidogenic Processing of Amyloid Precursor Protein Drives Stretch-Induced Disruption of Axonal Transport in hiPSC-Derived Neurons.J Neurosci. 2021 Dec 8;41(49):10034-10053. doi: 10.1523/JNEUROSCI.2553-20.2021. Epub 2021 Oct 18. J Neurosci. 2021. PMID: 34663629 Free PMC article.
-
Alzheimer's disease.Subcell Biochem. 2012;65:329-52. doi: 10.1007/978-94-007-5416-4_14. Subcell Biochem. 2012. PMID: 23225010 Review.
-
Subcellular trafficking of the amyloid precursor protein gene family and its pathogenic role in Alzheimer's disease.Neurodegener Dis. 2006;3(4-5):218-26. doi: 10.1159/000095259. Neurodegener Dis. 2006. PMID: 17047360 Review.
Cited by
-
Targeting increased levels of APP in Down syndrome: Posiphen-mediated reductions in APP and its products reverse endosomal phenotypes in the Ts65Dn mouse model.Alzheimers Dement. 2021 Feb;17(2):271-292. doi: 10.1002/alz.12185. Epub 2020 Sep 25. Alzheimers Dement. 2021. PMID: 32975365 Free PMC article.
-
Deregulation of signalling in genetic conditions affecting the lysosomal metabolism of cholesterol and galactosyl-sphingolipids.Neurobiol Dis. 2020 Dec;146:105142. doi: 10.1016/j.nbd.2020.105142. Epub 2020 Oct 17. Neurobiol Dis. 2020. PMID: 33080336 Free PMC article. Review.
-
UV irradiation accelerates amyloid precursor protein (APP) processing and disrupts APP axonal transport.J Neurosci. 2014 Feb 26;34(9):3320-39. doi: 10.1523/JNEUROSCI.1503-13.2014. J Neurosci. 2014. PMID: 24573290 Free PMC article.
-
Increased expression of reticulon 3 in neurons leads to reduced axonal transport of β site amyloid precursor protein-cleaving enzyme 1.J Biol Chem. 2013 Oct 18;288(42):30236-30245. doi: 10.1074/jbc.M113.480079. Epub 2013 Sep 4. J Biol Chem. 2013. PMID: 24005676 Free PMC article.
-
A Role for Drosophila Amyloid Precursor Protein in Retrograde Trafficking of L1-Type Cell Adhesion Molecule Neuroglian.Front Cell Neurosci. 2019 Jul 12;13:322. doi: 10.3389/fncel.2019.00322. eCollection 2019. Front Cell Neurosci. 2019. PMID: 31354437 Free PMC article.
References
-
- Papp H., Pakaski M., Kasa P. Presenilin-1 and the amyloid precursor protein are transported bidirectionally in the sciatic nerve of adult rat. Neurochem. Int. 2002;41:429–435. - PubMed
-
- Gunawardena S., Goldstein L.S. Disruption of axonal transport and neuronal viability by amyloid precursor protein mutations in Drosophila. Neuron. 2001;32:389–401. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
