

The histidine kinase PdhS controls cell cycle progression of the pathogenic alphaproteobacterium Brucella abortus
- PMID: 22843843
- PMCID: PMC3457221
- DOI: 10.1128/JB.00699-12
The histidine kinase PdhS controls cell cycle progression of the pathogenic alphaproteobacterium Brucella abortus
Abstract
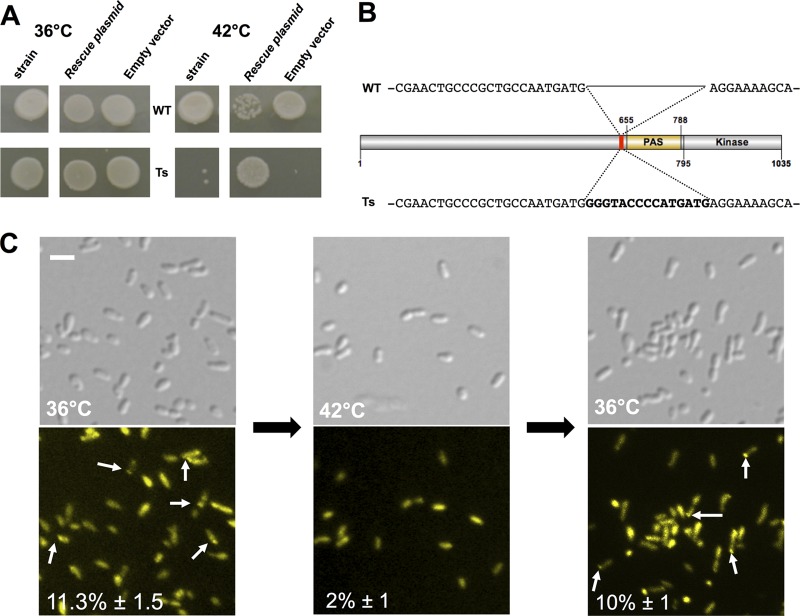
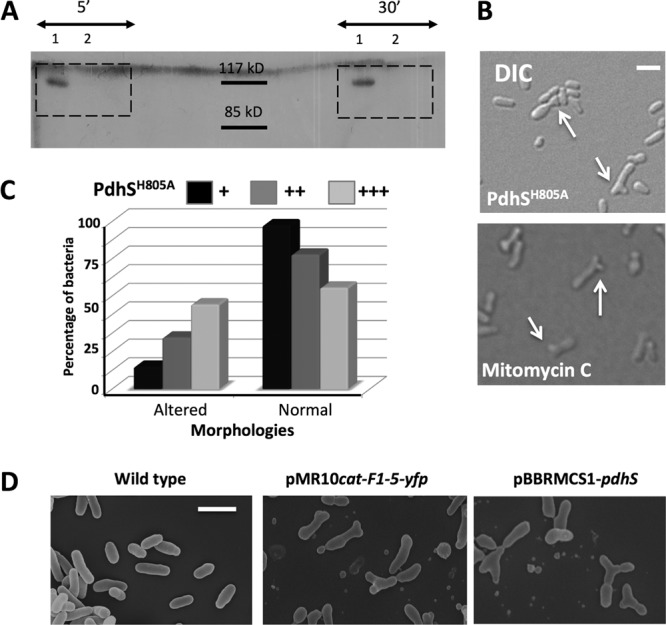
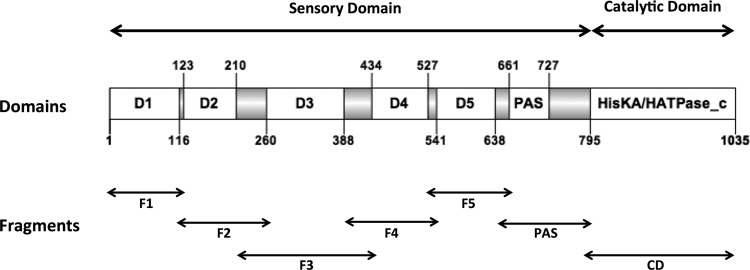
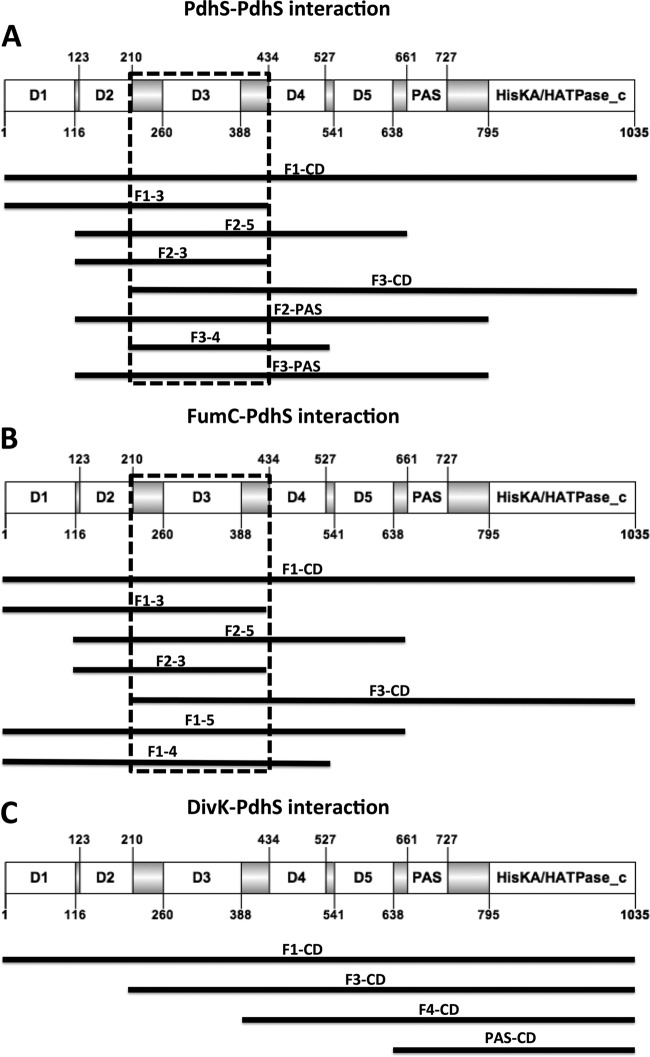
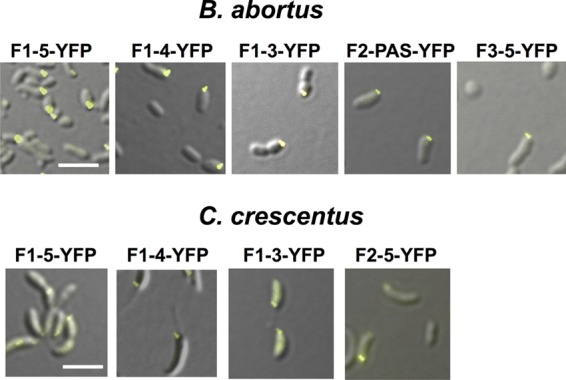
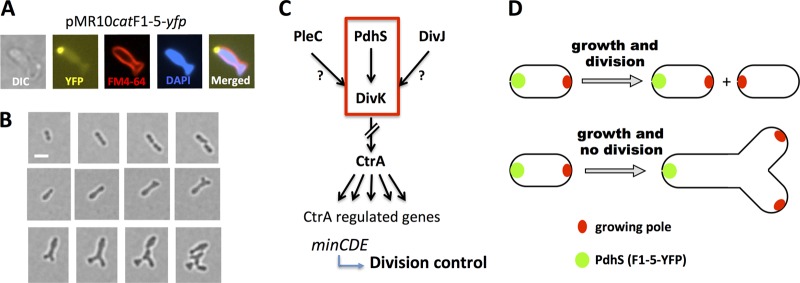
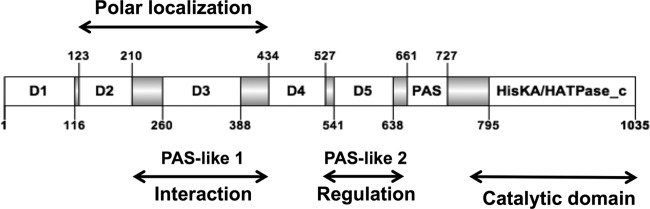
Bacterial differentiation is often associated with the asymmetric localization of regulatory proteins, such as histidine kinases. PdhS is an essential and polarly localized histidine kinase in the pathogenic alphaproteobacterium Brucella abortus. After cell division, PdhS is asymmetrically segregated between the two sibling cells, highlighting a differentiation event. However, the function(s) of PdhS in the B. abortus cell cycle remains unknown. We used an original approach, the pentapeptide scanning mutagenesis method, to generate a thermosensitive allele of pdhS. We report that a B. abortus strain carrying this pdhS allele displays growth arrest and an altered DivK-yellow fluorescent protein (YFP) polar localization at the restrictive temperature. Moreover, the production of a nonphosphorylatable PdhS protein or truncated PdhS proteins leads to dominant-negative effects by generating morphological defects consistent with the inhibition of cell division. In addition, we have used a domain mapping approach combined with yeast two-hybrid and fluorescence microscopy methods to better characterize the unusual PdhS sensory domain. We have identified a fragment of the PdhS sensory domain required for protein-protein interaction (amino acids [aa] 210 to 434), a fragment sufficient for polar localization (aa 1 to 434), and a fragment (aa 527 to 661) whose production in B. abortus correlates with the generation of cell shape alterations. The data support a model in which PdhS acts as an essential regulator of cell cycle progression in B. abortus and contribute to a better understanding of the differentiation program inherited by the two sibling cells.
Figures
References
-
- Bellefontaine AF, et al. 2002. Plasticity of a transcriptional regulation network among alpha-proteobacteria is supported by the identification of CtrA targets in Brucella abortus. Mol. Microbiol. 43:945–960 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
