

Functional epialleles at an endogenous human centromere
- PMID: 22847449
- PMCID: PMC3427087
- DOI: 10.1073/pnas.1203126109
Functional epialleles at an endogenous human centromere
Abstract
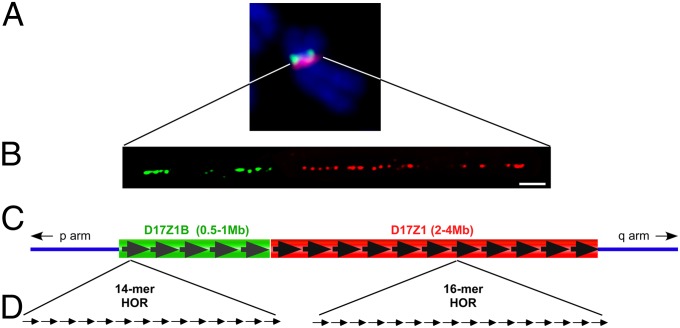
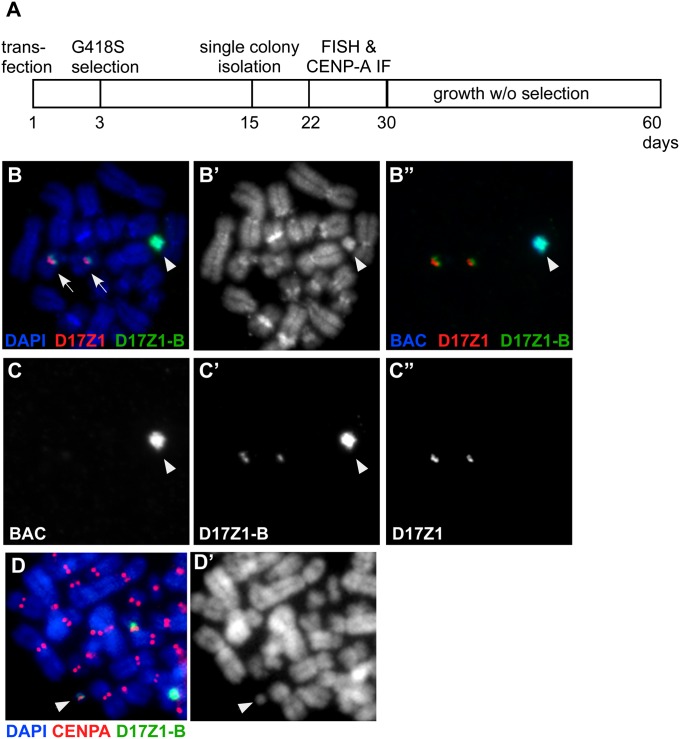
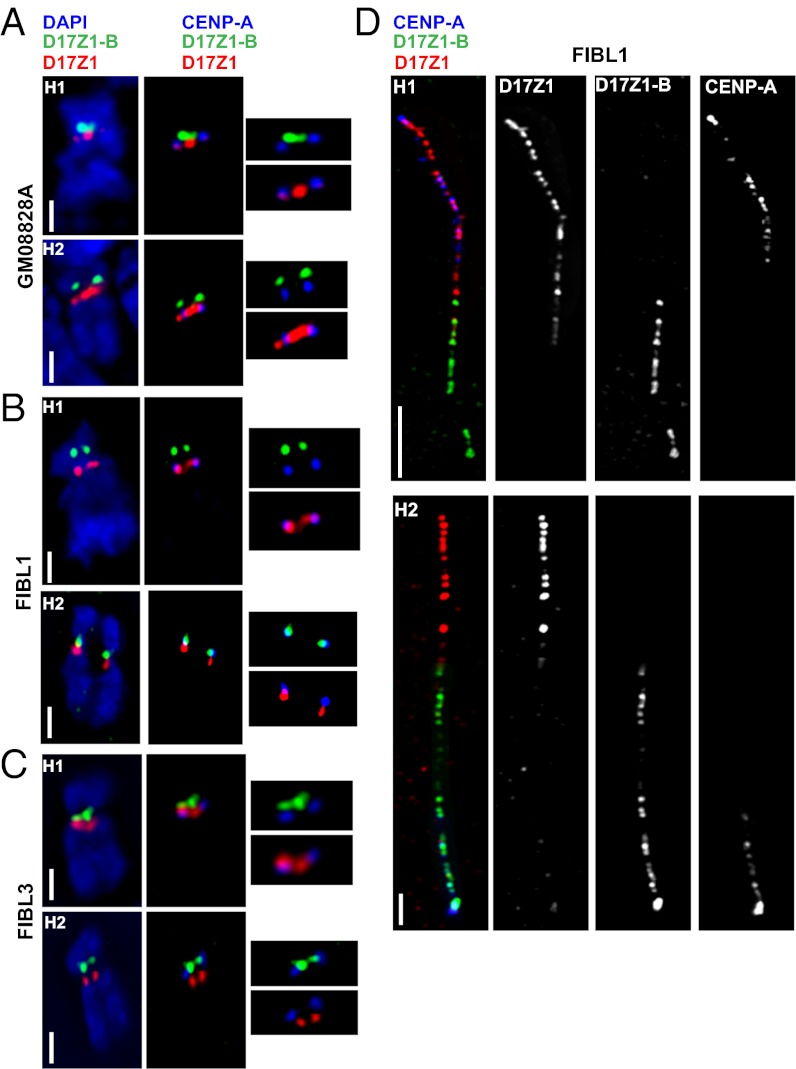
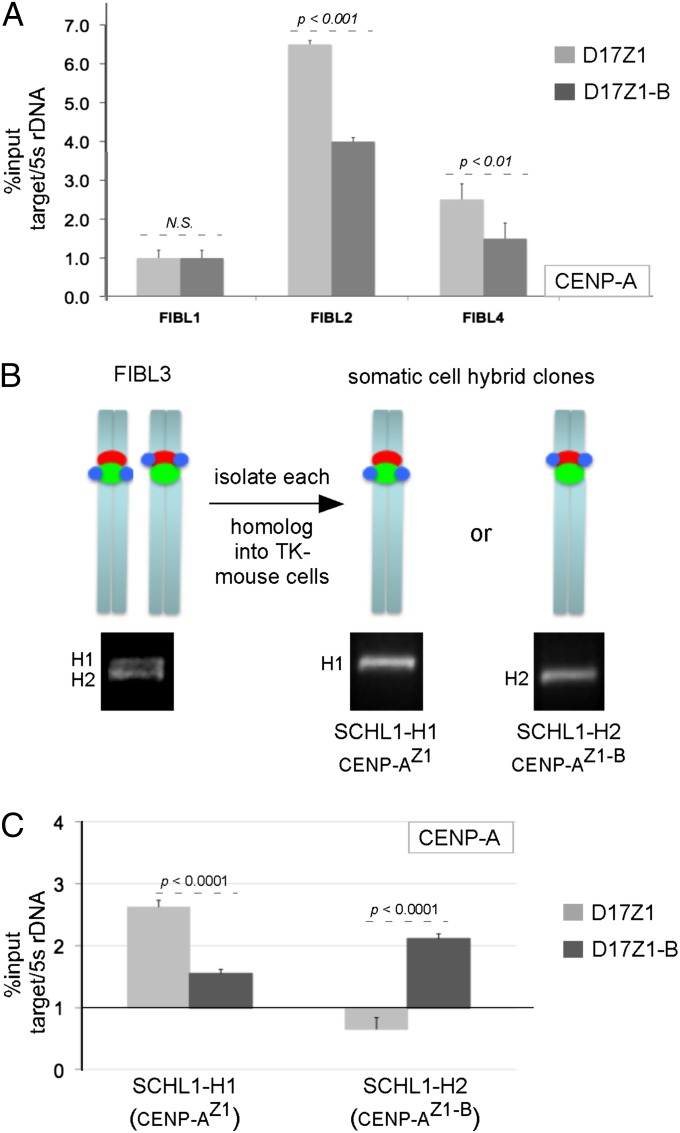
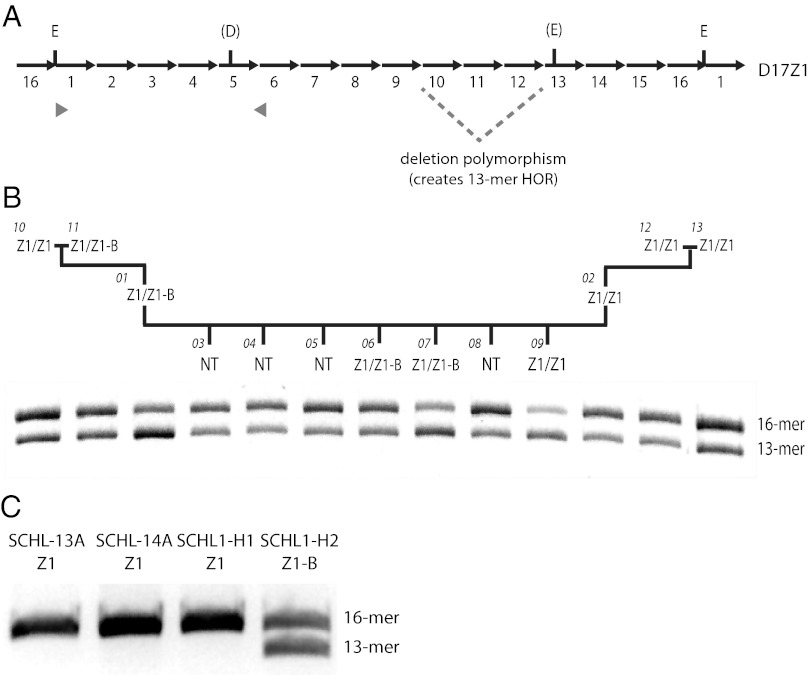
Human centromeres are defined by megabases of homogenous alpha-satellite DNA arrays that are packaged into specialized chromatin marked by the centromeric histone variant, centromeric protein A (CENP-A). Although most human chromosomes have a single higher-order repeat (HOR) array of alpha satellites, several chromosomes have more than one HOR array. Homo sapiens chromosome 17 (HSA17) has two juxtaposed HOR arrays, D17Z1 and D17Z1-B. Only D17Z1 has been linked to CENP-A chromatin assembly. Here, we use human artificial chromosome assembly assays to show that both D17Z1 and D17Z1-B can support de novo centromere assembly independently. We extend these in vitro studies and demonstrate, using immunostaining and chromatin analyses, that in human cells the centromere can be assembled at D17Z1 or D17Z1-B. Intriguingly, some humans are functional heterozygotes, meaning that CENP-A is located at a different HOR array on the two HSA17 homologs. The site of CENP-A assembly on HSA17 is stable and is transmitted through meiosis, as evidenced by inheritance of CENP-A location through multigenerational families. Differences in histone modifications are not linked clearly with active and inactive D17Z1 and D17Z1-B arrays; however, we detect a correlation between the presence of variant repeat units of D17Z1 and CENP-A assembly at the opposite array, D17Z1-B. Our studies reveal the presence of centromeric epialleles on an endogenous human chromosome and suggest genomic complexities underlying the mechanisms that determine centromere identity in humans.
Figures
References
-
- Palmer DK, O’Day K, Margolis RL. Biochemical analysis of CENP-A, a centromeric protein with histone-like properties. Prog Clin Biol Res. 1989;318:61–72. - PubMed
-
- Warburton PE, et al. Immunolocalization of CENP-A suggests a distinct nucleosome structure at the inner kinetochore plate of active centromeres. Curr Biol. 1997;7:901–904. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
