

Fluorescent in situ folding control for rapid optimization of cell-free membrane protein synthesis
- PMID: 22848743
- PMCID: PMC3407079
- DOI: 10.1371/journal.pone.0042186
Fluorescent in situ folding control for rapid optimization of cell-free membrane protein synthesis
Abstract
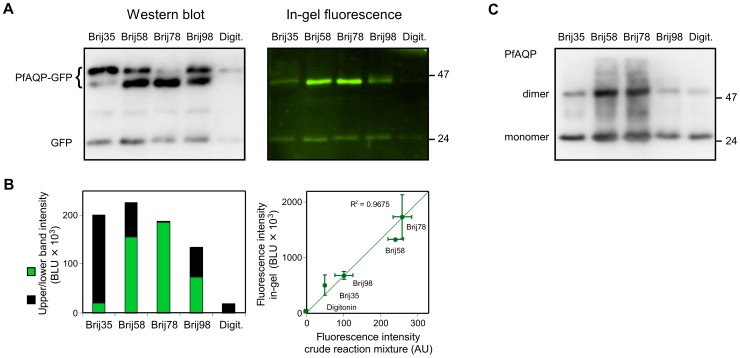
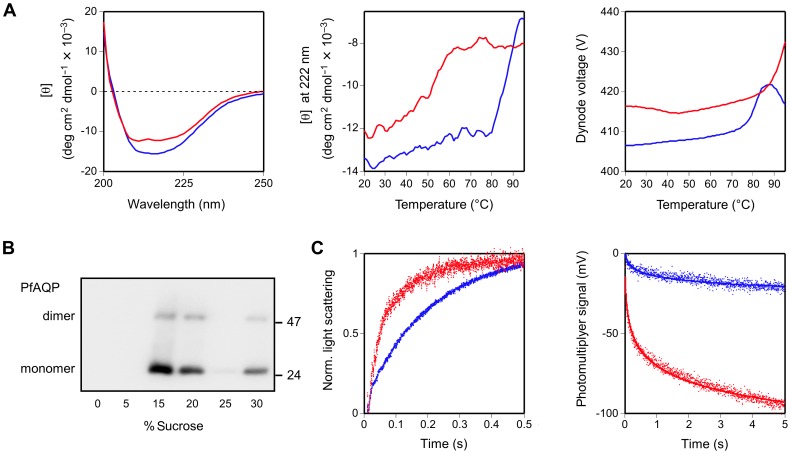
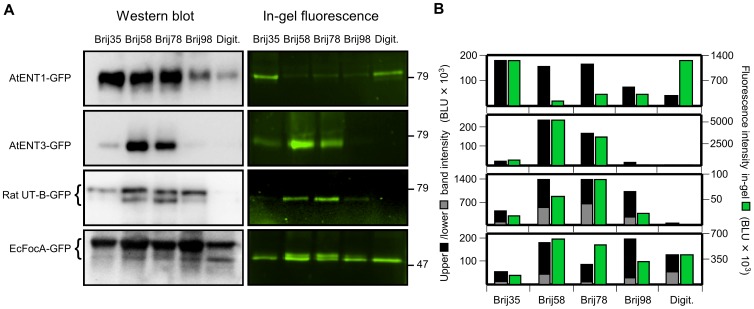
Cell-free synthesis is an open and powerful tool for high-yield protein production in small reaction volumes predestined for high-throughput structural and functional analysis. Membrane proteins require addition of detergents for solubilization, liposomes, or nanodiscs. Hence, the number of parameters to be tested is significantly higher than with soluble proteins. Optimization is commonly done with respect to protein yield, yet without knowledge of the protein folding status. This approach contains a large inherent risk of ending up with non-functional protein. We show that fluorophore formation in C-terminal fusions with green fluorescent protein (GFP) indicates the folding state of a membrane protein in situ, i.e. within the cell-free reaction mixture, as confirmed by circular dichroism (CD), proteoliposome reconstitution and functional assays. Quantification of protein yield and in-gel fluorescence intensity imply suitability of the method for membrane proteins of bacterial, protozoan, plant, and mammalian origin, representing vacuolar and plasma membrane localization, as well as intra- and extracellular positioning of the C-terminus. We conclude that GFP-fusions provide an extension to cell-free protein synthesis systems eliminating the need for experimental folding control and, thus, enabling rapid optimization towards membrane protein quality.
Conflict of interest statement
Figures
References
-
- Prasher DC, Eckenrode VK, Ward WW, Prendergast FG, Cormier MJ (1992) Primary structure of the Aequorea victoria green-fluorescent protein. Gene 111: 229–233. - PubMed
-
- Waldo GS, Standish BM, Berendzen J, Terwilliger TC (1999) Rapid protein-folding assay using green fluorescent protein. Nat Biotechnol 17: 691–695. - PubMed
-
- Drew D, Lerch M, Kunji E, Slotboom DJ, de Gier JW (2006) Optimization of membrane protein overexpression and purification using GFP fusions. Nat Methods 3: 303–313. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
