

Human RHOH deficiency causes T cell defects and susceptibility to EV-HPV infections
- PMID: 22850876
- PMCID: PMC3428089
- DOI: 10.1172/JCI62949
Human RHOH deficiency causes T cell defects and susceptibility to EV-HPV infections
Abstract
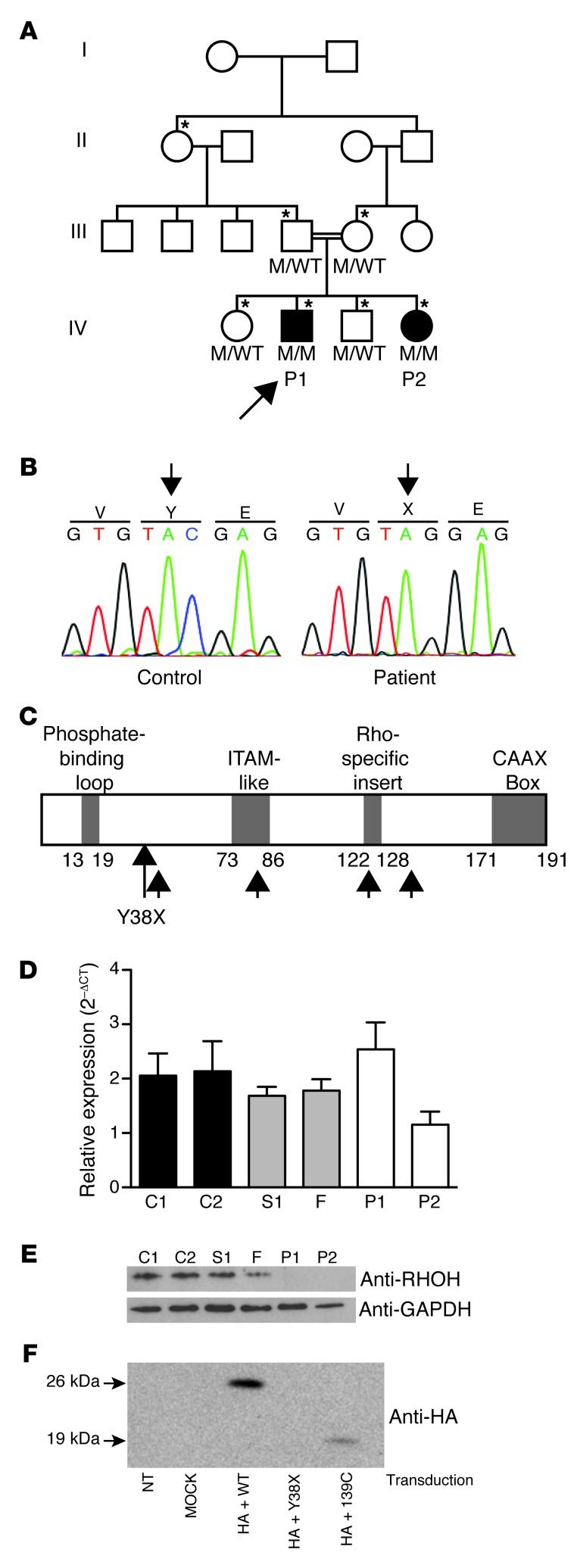
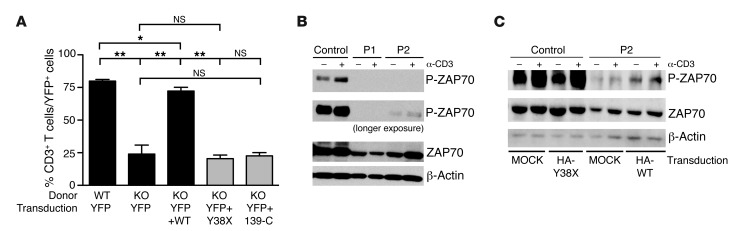
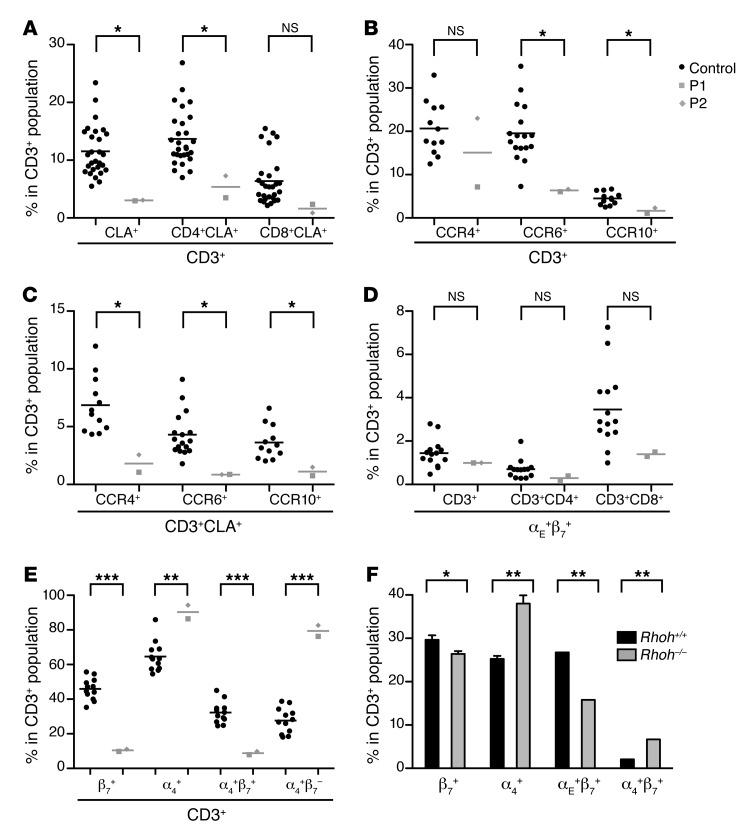
Epidermodysplasia verruciformis (EV) is a rare genetic disorder characterized by increased susceptibility to specific human papillomaviruses, the betapapillomaviruses. These EV-HPVs cause warts and increase the risk of skin carcinomas in otherwise healthy individuals. Inactivating mutations in epidermodysplasia verruciformis 1 (EVER1) or EVER2 have been identified in most, but not all, patients with autosomal recessive EV. We found that 2 young adult siblings presenting with T cell deficiency and various infectious diseases, including persistent EV-HPV infections, were homozygous for a mutation creating a stop codon in the ras homolog gene family member H (RHOH) gene. RHOH encodes an atypical Rho GTPase expressed predominantly in hematopoietic cells. Patients' circulating T cells contained predominantly effector memory T cells, which displayed impaired TCR signaling. Additionally, very few circulating T cells expressed the β7 integrin subunit, which homes T cells to specific tissues. Similarly, Rhoh-null mice exhibited a severe overall T cell defect and abnormally small numbers of circulating β7-positive cells. Expression of the WT, but not of the mutated RHOH, allele in Rhoh-/- hematopoietic stem cells corrected the T cell lymphopenia in mice after bone marrow transplantation. We conclude that RHOH deficiency leads to T cell defects and persistent EV-HPV infections, suggesting that T cells play a role in the pathogenesis of chronic EV-HPV infections.
Figures
References
-
- Lutzner MA. Epidermodysplasia verruciformis. An autosomal recessive disease characterized by viral warts and skin cancer. A model for viral oncogenesis. Bull Cancer. 1978;65(2):169–182. - PubMed
-
- Orth G. Host defenses against human papillomaviruses: lessons from epidermodysplasia verruciformis. Curr Top Microbiol Immunol. 2008;321:59–83. - PubMed
-
- Cockayne E. Inherited Abnormalities Of The Skin And Its Appendages. London, United Kingdom: Oxford University Press; 1933.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
