

Enzymatic engineering of polysialic acid on cells in vitro and in vivo using a purified bacterial polysialyltransferase
- PMID: 22851175
- PMCID: PMC3463313
- DOI: 10.1074/jbc.M112.377614
Enzymatic engineering of polysialic acid on cells in vitro and in vivo using a purified bacterial polysialyltransferase
Abstract
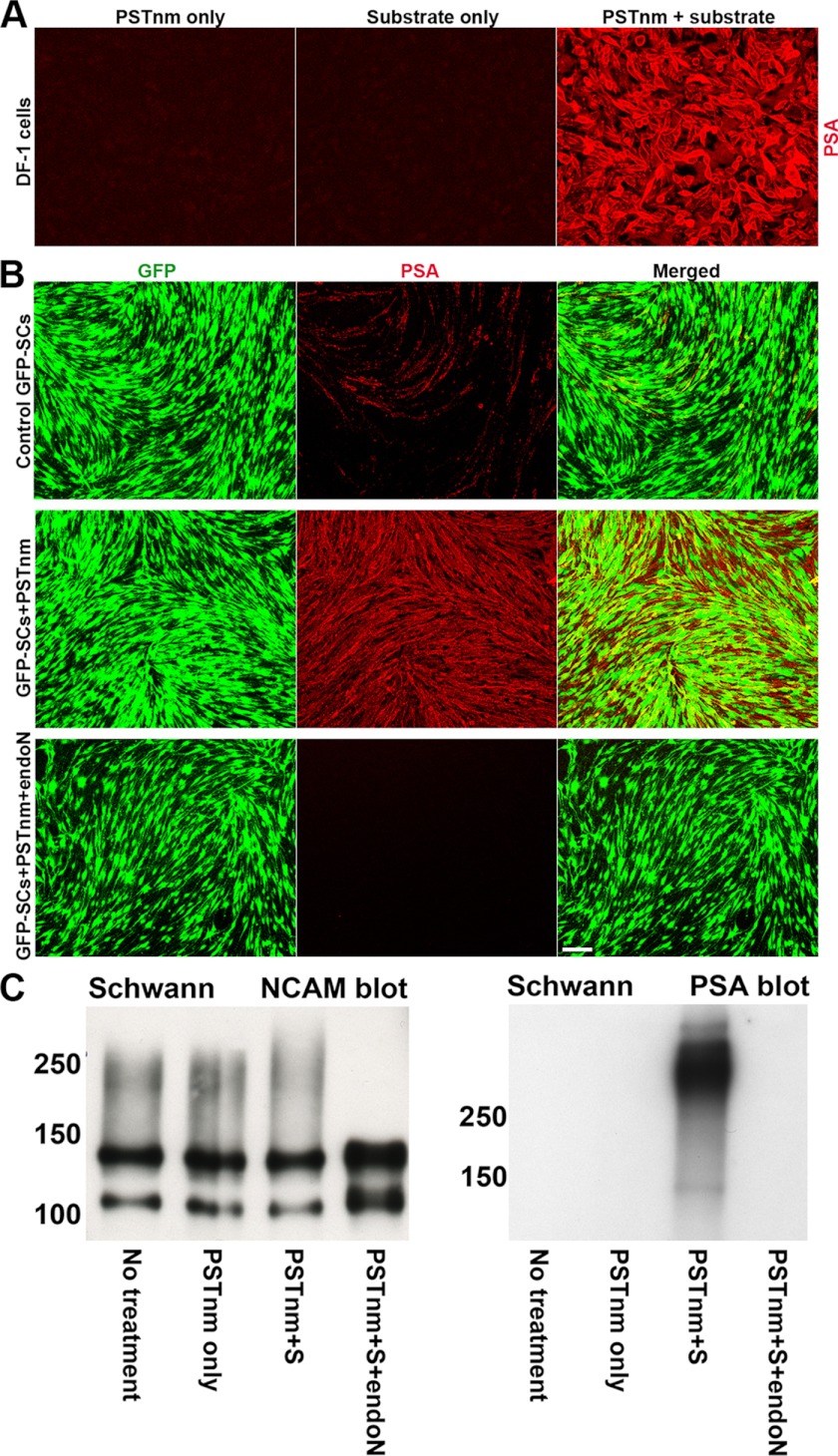
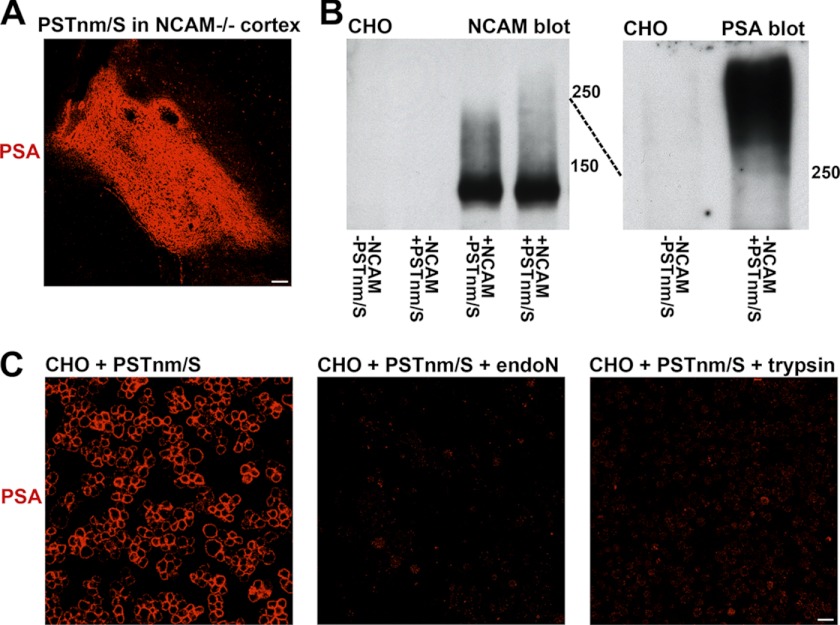
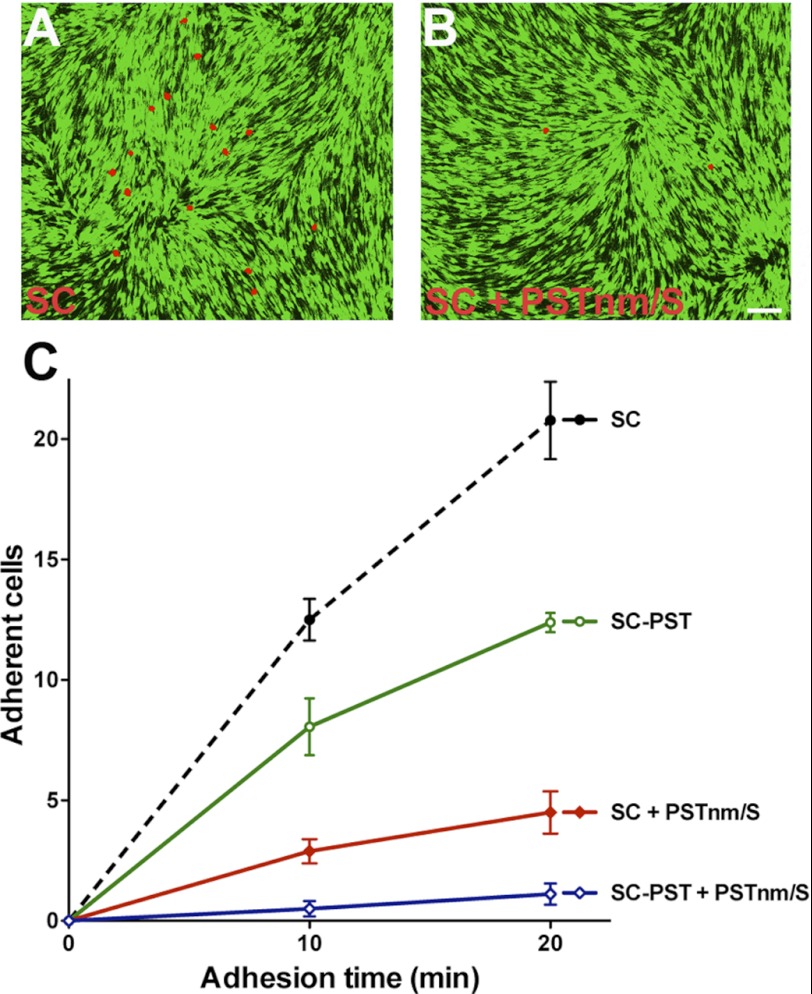
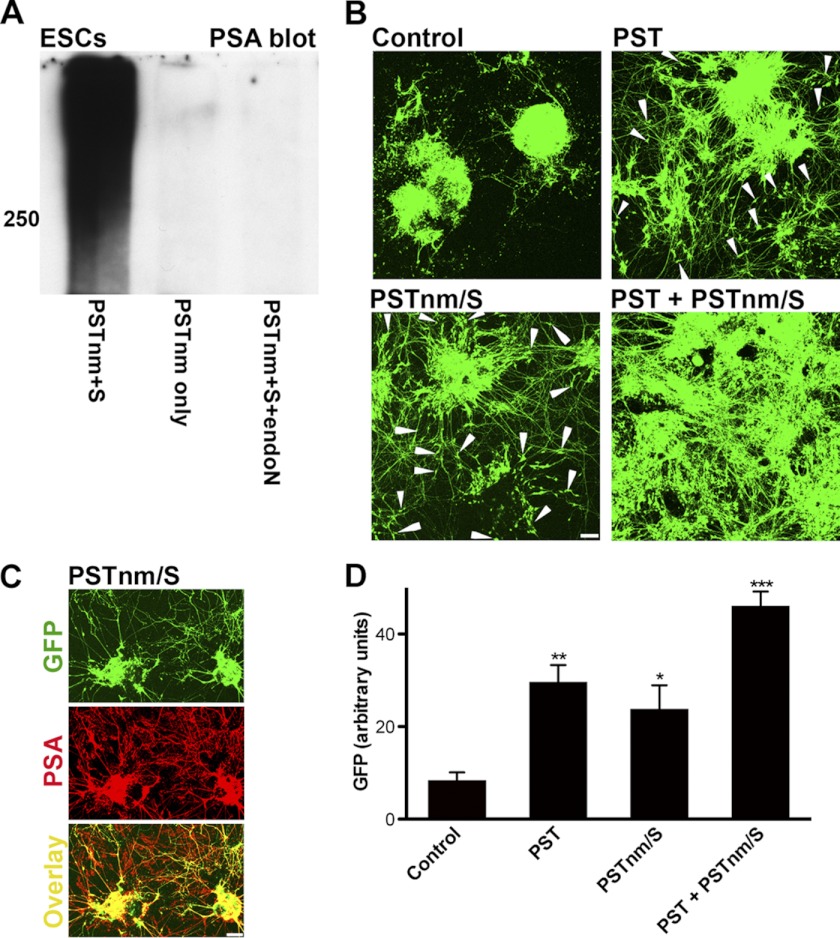
In vertebrates, polysialic acid (PSA) is typically added to the neural cell adhesion molecule (NCAM) in the Golgi by PST or STX polysialyltransferase. PSA promotes plasticity, and its enhanced expression by viral delivery of the PST or STX gene has been shown to promote cellular processes that are useful for repair of the injured adult nervous system. Here we demonstrate a new strategy for PSA induction on cells involving addition of a purified polysialyltransferase from Neisseria meningitidis (PST(Nm)) to the extracellular environment. In the presence of its donor substrate (CMP-Neu5Ac), PST(Nm) synthesized PSA directly on surfaces of various cell types in culture, including Chinese hamster ovary cells, chicken DF1 fibroblasts, primary rat Schwann cells, and mouse embryonic stem cells. Similarly, injection of PST(Nm) and donor in vivo was able to produce PSA in different adult brain regions, including the cerebral cortex, striatum, and spinal cord. PSA synthesis by PST(Nm) requires the presence of the donor CMP-Neu5Ac, and the product could be degraded by the PSA-specific endoneuraminidase-N. Although PST(Nm) was able to add PSA to NCAM, most of its product was attached to other cell surface proteins. Nevertheless, the PST(Nm)-induced PSA displayed the ability to attenuate cell adhesion, promote neurite outgrowth, and enhance cell migration as has been reported for endogenous PSA-NCAM. Polysialylation by PST(Nm) occurred in vivo in less than 2.5 h, persisted in tissues, and then decreased within a few weeks. Together these characteristics suggest that a PST(Nm)-based approach may provide a valuable alternative to PST gene therapy.
Figures


References
-
- Eckhardt M., Mühlenhoff M., Bethe A., Koopman J., Frosch M., Gerardy-Schahn R. (1995) Molecular characterization of eukaryotic polysialyltransferase-1. Nature 373, 715–718 - PubMed
-
- Johnson C. P., Fujimoto I., Rutishauser U., Leckband D. E. (2005) Direct evidence that neural cell adhesion molecule (NCAM) polysialylation increases intermembrane repulsion and abrogates adhesion. J. Biol. Chem. 280, 137–145 - PubMed
-
- Fujimoto I., Bruses J. L., Rutishauser U. (2001) Regulation of cell adhesion by polysialic acid: effects on cadherin, immunoglobulin cell adhesion molecule, and integrin function and independence from neural cell adhesion molecule binding or signaling activity. J. Biol. Chem. 276, 31745–31751 - PubMed
-
- Rutishauser U. (2008) Polysialic acid in the plasticity of the developing and adult vertebrate nervous system. Nat. Rev. Neurosci. 9, 26–35 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
